Startseite - Impressum - Datenschutzerklärung - News - Ich diene dem, der hört - https://offenunddirekt.de - Mein Gratis-Coaching für Dich - Kraft tanken - Spirituelles - Kunst - Technik - Literatur - Kultur - Politik - Architektur - Spaß - Psychologie - Spiele - Fotos + Animationen - Gästebuch - Persönliches - Blog - Die Stimme Gottes - mein Youtube Kanal - erstes Rätsel - Renaissance Forum - Chat - Quiz - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - Zufallsspiel - letztes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Ernst Haeckel - Die Natürliche Schöpfungsgeschichte 1868
Achtzehnter Vortrag.
Stammbaum und Geschichte des Thierreichs.
II. Stammbaum und Geschichte der Wirbelthiere.
(Hierzu Taf. VI und VII.)

Meine Herren! Unter den natürlichen Hauptgruppen der Organismen, welche wir wegen der Blutsverwandtschaft aller darin vereinigten Arten als Stämme oder Phylen bezeichnen, ist keine einzige von so hervorragender und überwiegender Bedeutung, als der Stamm der Wirbelthiere. Denn nach dem übereinstimmenden Urtheil aller Zoologen ist auch der Mensch ein Glied dieses Stammes, und kann seiner ganzen Organisation und Entwickelung nach unmöglich von den übrigen Wirbelthieren getrennt werden. Wie wir aber aus der individuellen Entwickelungsgeschichte des Menschen schon früher die unbestreitbare Thatsache erkannt haben, daß derselbe in seiner Entwickelung aus dem Ei anfänglich nicht von den übrigen Wirbelthieren, und namentlich den Säugethieren verschieden ist, so müssen wir nothwendig mit Beziehung auf seine paläontologische Entwickelungsgeschichte schließen, daß das Menschengeschlecht sich historisch wirklich aus niederen Wirbelthieren entwickelt hat, und daß dasselbe zunächst von den Säugethieren abstammt. Dieser Umstand allein schon (abgesehen von dem vielseitigen höheren Interesse, das auch in anderer Beziehung die Wirbelthiere vor den übrigen Organismen in Anspruch nehmen) wird es rechtfertigen, daß wir den Stammbaum der Wirbelthiere und dessen Ausdruck, das natürliche System, hier besonders genau untersuchen.
Die Bezeichnung Wirbelthiere (Vertebrata) rührt, wie ich schon im letzten Vortrage erwähnte, von dem großen Lamarck her, welcher zuerst gegen Ende des vorigen Jahrhunderts unter diesem Namen die vier oberen Thierklassen Linné's zusammenfaßte: die Säugethiere, Vögel, Amphibien und Fische. Die beiden niederen Klassen Linné's, die Insecten und Würmer, stellte Lamarck den Wirbelthieren gegenüber als Wirbellose (Invertebrata, später auch Evertebrata genannt). Die Eintheilung der Wirbelthiere in die vier genannten Klassen wurde auch von Cuvier und seinen Nachfolgern, und in Folge dessen von vielen Zoologen noch bis auf die Gegenwart festgehalten. Aber schon 1822 erkannte der ausgezeichnete Anatom Blainville aus der vergleichenden Anatomie, und fast gleichzeitig unser großer Embryologe Bär aus der Ontogenie der Wirbelthiere, daß Linné's Klasse der Amphibien eine unnatürliche Vereinigung von zwei ganz verschiedenen Klassen sei. Diese beiden Klassen hatte schon 1820 Merrem als zwei Hauptgruppen der Amphibien unter den Namen Pholidoten und Batrachier getrennt. Die Batrachier, welche heutzutage gewöhnlich als Amphibien (im engeren Sinne!) bezeichnet werden, umfassen die Frösche, Salamander, Kiemenmolche, Cäcilien und die ausgestorbenen Labyrinthodonten. Sie schließen sich in ihrer ganzen Organisation eng an die Fische an. Die Pholodoten oder Reptilien dagegen sind viel näher den Vögeln verwandt. Es gehören dahin die Eidechsen, Schlangen, Krocodile und Schildkröten, und die vielgestaltige Formengruppe der mesolithischen Drachen, Seedrachen, Flugeidechsen u. s. .w
Im Anschluß an diese naturgemäße Scheidung der Amphibien in zwei Klassen theilte man nun den ganzen Stamm der Wirbelthiere in zwei Hauptgruppen. Die erste Hauptgruppe, die Fische und Amphibien, athmen entweder zeitlebens oder doch in der Jugend durch Kiemen, und werden daher als Kiemenwirbelthiere bezeichnet (Branchiata oder Anallontoidia). Die zweite Hauptgruppe dagegen, Reptilien, Vögel, und Säugethiere, athmen zu keiner Zeit ihres Lebens durch Kiemen, sondern ausschließlich durch Lungen, und heißen deshalb auch passend kiemenlose oder Lungenwirbelthiere (Ebranchiata oder Allantoidia). So richtig diese Unterscheidung auch ist, so können wir doch bei derselben nicht stehen bleiben, wenn wir zu einem wahren natürlichen System des Wirbelthierstammes, und zu einem naturgemäßen Verständniß seines Stammbaums gelangen wollen. Vielmehr müssen wir dann, wie ich vor zwei Jahren in meiner generellen Morphologie gezeigt habe, noch drei weitere Wirbelthierklassen unterscheiden, indem wir die bisherige Fischklasse in vier verschiedene Klassen auflösen (Gen. Morph. II. Bd., Taf. VII, S. CXVI-CLX).
Die erste und niederste von diesen Klassen wird durch die Rohrherzen (Leptocardia) oder Schädellosen (Acrania) gebildet, von denen heutzutage nur noch ein einziger Repräsentant lebt, das merkwürdige Lanzetthierchen (Amphioxus lanceolatus). Als zweite Klase schließen sich an diese zunächst die Unpaarnasen (Monorrhina) oder Rundmäuler (Cyclostoma) an, zu denen die Inger (Myxinoiden) und die Lampreten (Petromyzonten) gehören. Die dritte Klasse erst würde die echten Fische (Pisces) bilden und an diese würden sich als vierte Klasse die Lurchfische (Dipneusta) anschließen: Uebergangsformen zu den Amphibien. Durch diese Unterscheidung, welche, wie Sie gleich sehen werden, für die Genealogie der Wirbelthiere sehr wichtig ist, wird die ursprüngliche Vierzahl der Wirbelthierklassen auf das Doppelte gesteigert.
Diese acht Klassen der Wirbelthiere sind aber keineswegs von gleichem genealogischem Werthe. Vielmehr müssen wir dieselben in der Weise, wie es Ihnen bereits die systematische Uebersicht auf S. 393 zeigte, auf die vier verschiedenen Hauptklassen vertheilen. Zunächst können wir die drei höchsten Klassen, die Säugethiere, Vögel und Schleicher als eine natürliche Hauptklasse unter dem Namen der Amnionthiere (Amniota) zusammenfassen. Diesen stellen sich natürgemäß als eine zweite Hauptklasse die Amionlosen (Anamnia) gegenüber, nämlich die drei Klassen der Lurche, Lurchfische und Fische. Die genannten sechs Klassen, sowohl die Amnionlosen als die Amnionthiere, stimmen unter sich in zahlreichen Merkmalen überein, durch welche sie sich von den beiden niedersten Klassen (den Unpaarnasen und Rohrherzen) unterscheiden. Wir können sie daher in der natürlichen Hauptgruppe der Paarnasen (Amphirrhina) vereinigen. Endlich sind diese Paarnasen wiederum viel näher den Rundmäulern oder Unpaarnasen, als den Schädellosen oder Rohrherzen verwandt. Wir können daher mit vollen Rechte die Paarnasen mit den Unpaarnasen in einer obersten Hauptgruppe zusammenstellen und diese als Centralherzen (Pachycardia) oder Schädelthiere (Craniota) der einzigen Klasse der Rohrherzen oder Schädellosen gegenüberstellen. Das systematische Verhältniß dieser Gruppen zu einander wird Ihnen durch folgende Uebersicht klar werden.
{ Tabelle Seite 436 unten }

Auf der niedrigsten Organisationsstufe von allen uns bekannten Wirbelthieren steht der einzige noch lebende Vertreter der ersten Klasse, das Lanzetfischchen oder Lanzetthierchen (Amphioxus lanceolatus). Dieses höchst interessante und wichtige Thierchen, welches über die älteren Wurzeln unsres Stammbaumes ein überraschendes Licht verbreitet, ist offenbar der letzte Mohikaner, der letzte überlebende Repräsentant einer formenreichen niederen Wirbelthierklasse, welche während der Primordialzeit sehr entwickelt war, uns aber leider wegen des Mangels aller festen Skelettheile gar keine versteinerten Reste hinterlassen konnte. Das kleine Lanzetfischchen lebt heut noch weiterverbreitet in verschiedenen Meeren, z. b. in der Ostsee, Nordsee, im Mittelmeere, gewöhnlich auf flachem Strande im Sand vergraben. Der Körper hat, wie der Name sagt, die Gestalt eines schmalen, an beiden Enden zugespitzten, lanzettförmigen Blattes. Erwachsen ist dasselbe etwa zwei Zoll lang, und röthlich schimmernd, halb durchsichtig. Aeußerlich hat das Lanzetthierchen so wenig Aehnlichkeit mit einem Wirbelthier, daß sein erster Entdecker, Pallas, es für eine unvollkommene Nacktschnecke hielt. Beine besitzt es nicht, und ebenso wenig Kopf, Schädel und Gehirn. Das vordere Körperende ist äußerlich von dem hinteren fast nur durch die Mundöffnung zu unterscheiden. Aber dennoch besitzt der Amphioxus in seinem inneren Bau die wichtigsten Merkmale, durch welche sich alle Wirbelthiere von allen Wirbellosen unterscheiden, vor allen den Rückenstrang und das Rückenmark.
Der Rückenstrang (Chorda dorsalis) ist ein cylindrischer, vorn und hinten zugespitzter, grader Knorpelstab, welcher die centrale Axe des inneren Skelets, und die Grundlage der Wirbelsäule bildet. Umittelbar über diesem Rückenstrang, auf der Rückenseite desselben, liegt das Rückenmark (Medulla spinalis), ebenfalls ursprünglich ein grader, vorn und hinten zugespitzter, inwendig aber hohler Strang, welcher das Hauptstück und Centrum des Nervensystems bei allen Wirbelthieren bildet (Vergl. oben S. 247, 248). Bei allen Wirbelthieren ohne Ausnahme, auch den Menschen mit inbegriffen, werden diese wichtigsten Körpertheile während der embryonalen Entwickelung aus dem Ei ursprünglich in derselben einfachsten Form angelegt, welche sie beim Amphioxus zeitlebens behalten. Erst später entwickelt sich durch die Auftreibung des vorderen Endes aus dem Rückenmark das Gehirn, und aus dem Rückenstrang der das Gehirn umschließende Schädel.
Da bei dem Amphioxus diese beiden wichtigen Organe gar nicht zur Entwickelung gelangen, so können wir die durch ihn vertretenen Thierklasse mit Recht als Schädellose (Acrania) bezeichnen, im Gegensatz zu allen übrigen, den Schädelthieren (Craniota). Gewöhnlich werden die Schädellosen Rohrherzen oder Röhrenherzen (Leptocardia) genannt, weil ein centralisirtes Herz noch fehlt, und das Blut durch die Zusammenziehungen der röhrenförmigen Blutgefäße selbst im Körper umhergetrieben wird. Die Schädelthiere, welche dagegen ein centralisirtes, beutelförmiges Herz besitzen, müßten dann im Gegensatz dazu Beutelherzen oder Centralherzen (Pachycardia) genannt werden.
Offenbar haben sich die Schädelthiere oder Centralherzen erst in späterer Primordialzeit aus Schädellosen oder Rohrherzen, welche dem Amphioxus nahe standen, allmählich entwickelt. Darüber läßt uns die Ontogenie der Schädelthere nicht in Zweifel. Wo stammen nun aber diese Schädellosen selbst her? Auf diese wichtige Frage hat uns, wie ich schon im letzten Vortrage erwähnte, erst die jüngste Zeit eine höchst überraschende Antwort gegenben. Aus den 1867 veröffentlichten Untersuchungen von Kowalewski über die individuelle Entwickelung des Amphioxus und der festsitzenden Seescheiden (Ascidiae) [aus der Klasse der Mantelthiere (Tunicata)] hat sich ergeben, daß die Ontogenie dieser beiden ganz verschiedenen Thierformen in ihrer ersten Jugend merkwürdig übereinstimmt. Die frei umherschwimmenden Larven der Ascidien entwickeln die unzweifelhafte Anlage zum Rückenmark und zum Rückenstrang, und zwar ganz in derselben Weise, wie der Amphioxus. Allerdings bilden sie diese wichtigsten Organe des Wirbelthierkörpers späterhin nicht weiter aus. Vielmehr gehen sie eine rückschreitende Verwandlung ein, setzen sich auf dem Meeresboden fest, und wachsen zu unförmigen Klumpen aus, in denen man kaum noch bei äußerer Betrachtung ein Thier vermuthet. Allein das Rückenmark, als die Anlage des Centralnervensystems, und der Rückenstrang, als die erste Grundlage der Wirbelsäule, sind so wichtige, den Wirbelthieren so ausschließlich eigenthümliche Organe, daß wir daraus sicher auf die wirkliche Blutsverwandtschaft der Wirbelthiere mit den Mantelthieren schließen können. Natürlich wollen wir damit nicht sagen, daß die Wirbelthiere von den Mantelthieren abstammen, sondern nur, daß beide Gruppen aus gemeinsamer Wurzel entsprossen sind, und daß die Mantelthiere von allen Wirbellosen diejenigen sind, welche die nächste Blutsverwandtschaft zu den Wirbelthieren besitzen. Offenbar haben sich während der Primordialzeit die echten Wirbelthiere (und zwar zunächst die Schädellosen) aus einer Würmergruppe fortschreitend entwickelt, aus welcher nach einer anderen rückschreitenden Richtung hin, die degenerirten Mantelthiere hervorgingen.
Aus den Schädellosen oder Rohrherzen hat sich zunächst eine zweite niedere Klasse von Wirbelthieren entwickelt, welche noch tief unter den Fischen steht, und welche in der Gegenwart nur durch die Inger (Myxinoiden) und Lampreten (Petromyzonten) vertreten wird. Auch diese Klasse konnte wegen des Mangels aller festen Körpertheile leider eben so wenig als die Schädellosen versteinerte Reste hinterlassen. Aus ihrer ganzen Organisation und Ontogenie geht aber deutlich hervor, daß sie eine sehr wichtige Mittelstufe zwischen den Schädellosen und den Fischen darstellt, und daß die wenigen noch lebenden Glieder derselben nur die letzten überlebenden Reste von einer gegen Ende der Primordialzeit vermuthlich reich entwickelten Thiergruppe sind. Wegen des kreisrunden, zum Saugen verwendeten Maules, das die Inger und Lampreten besitzen, wird die ganze Klasse gewöhnlich Rundmäuler (Cyclostoma) genannt. Bezeichnender noch ist der Name Unpaarnasen (Monorrhina). Denn alle Cyclostomen besitzen ein einfaches sunpaares Nasenrohr, während bei allen übrigen Wirbelthieren (wieder mit Ausnahme des Amphioxus) die Nase aus zwei paarigen Seitenhälften, einer rechten und linken Nase besteht. Wir konnten deshalb diese letztren (Anamnien und Amnioten) auch als Paarnasen (Amphirrhina) zusammenfassen.
Auch abgesehen von der eigenthümlichen Nasenbildung unterscheiden sich die Unpaarnasen von den Paarnasen noch durch viele andere Eigenthümlichkeiten. So fehlt ihnen namentlich ganz das wichtige sympathische Nervennetz der letzteren. Ebenso wenig besitzen sie die Milz und die Bauchspeicheldrüse der Paarnasen. Von der Schwimmblase und den beiden Beinpaaren, welche bei allen Paarnasen wenigstens in der Anlage vorhanden sind, fehlt den Unpaarnasen (ebenso wie den Schädellosen) noch jede Spur. Es ist daher gewiß ganz gerechtfertigt, wenn wir sowohl die Monorrhinen als die Schädellosen gänzlich von den Fschen trennen, mit denen sie bis jetzt in herkömmlicher, aber irrthümlicher Weise vereinigt waren.
Die erste genauere Kenntniß der Monorrhinen oder Cyclostomen verdanken wir dem großen Berliner Zoologen Johannes Müller, dessen klassisches Werk über die "vergleichende Anatomie der Myxinoiden" die Grundlage unserer neueren Ansichten über den Bau der Wirbelthiere bildet. Er unterschied unter den Cyclostomen zwei verschiedene Gruppen, welche wir den Werth von Unterklassen geben können. Die erste Unterklasse sind die Inger oder Schleimfische (Hyperotreta oder Myxinoida). Sie leben im Meere schmarotzend auf anderen Fischen, in deren Haut sie sich einbohren (Myxine, Bdellostoma). Im Gehörorgan besitzen sie nur einen Ringkanal, und ihr unpaares Nasenrohr durchbohrt den Gaumen. Höher entwickelt ist die zweite Unterklasse, die Lampreten oder Pricken (Hyperoartia oder Petromyzontia). Hierher gehören die allbekannten Flußpricken oder Neunaugen unserer Flüsse (Petromyzon fluviatilis), deren Bekantschaft Sie wohl Alle im marinirten Zustande schon gemacht haben. Im Meere werden dieselben durch die mehrmals größeren Seepricken oder die eigentlichen Lampreten (Petromyzon marinus) vertreten. Bei diesen Unpaarnasen durchbohrt das Nasenrohr den Gaumen nicht, und im Gehörorgan finden sich zwei Ringcanäle.
{ Tabelle Seite 441 }

Alle Wirbelthiere, welche jetzt noch leben, mit Ausnahme der eben betrachteten Monorrhinen und des Amphioxus, gehören zu derjenigen Hauptgruppe, welche wir als Paarnasen (Amphirrhina) bezeichneten. Alle diese Thiere besitzen (trotz der großen Mannichfaltigkeit in ihrer sonstigen Bildung) eine aus zwei paarigen Seitenhälften bestehende Nase, ein sympatisches Nervennetz, drei Ringcanäle im Gehörorgan, ferner eine blasenförmige Ausstülpung des Schlundes, welche sich bei den Fischen zur Schwimmblase, bei den übrigen Paarnasen zur Lunge entwickelt hat. Endlich ist ursprünglich bei allen Paarnasen die Anlage von zwei paar Extremitäten oder Gliedmaßen vorhanden, ein paar Vorderbeine oder Brustflossen, und ein paar Hinterbeine oder Bauchflossen. Allerdings ist bisweilen das eine Beinpaar (z. b. bei den Aalen und Walfischen) oder beide Beinpaare (z. B. bei den Caecilien und Schlangen) verkümmert oder gänzlich verloren gegangen; aber selbst in diesen Fällen ist wenigstens eine Spur ihrer ursprünglichen Anlage in früher Embryonalzeit zu finden, oder es bleiben unnütze Reste derselben als rudimentäre Organe durch das ganze Leben bestehen (Vergl. oben S. 11).
Aus allen diesen wichtigen Anzeichen können wir mit voller Sicherheit schließen, daß sämmtliche Paarnasen von einer einzigen gemeinschaftlichen Stammform abstammen, welche während der Primordialzeit direct oder indirect sich aus den Monorrhinen entwickelt hatte. Diese Stammform muß die eben angeführten Organe, namentlich auch die Anlage zur Schwimmblase und zu zwei Beinpaaren oder Flossenpaaren besessen haben. Von allen jetzt lebenden Paarnasen stehen offenbar die niedersten Formen der Haifische dieser längst ausgestorbenen, unbekannten, hypothetischen Stammform, welche wir als Stammpaarnasen oder Proselachier bezeichnen können, am nächsten (Taf. VI, 11). Wir dürfen daher die Gruppe der Urfische oder Selachier, in deren Rahmen diese Proselachier vermuthlich hineingepaßt haben, als die Stammgruppe nicht allein der Fischklasse, sondern für die ganze Hauptklasse der Paarnasen betrachten.
Die Klasse der Fische (Pisces), mit welcher wir demgemäß die Reihe der Paarnasen beginnen, unterscheidet sich von den übrigen fünf Klassen dieser Reihe vorzüglich dadurch, daß die Schwimmblase niemals zur Lunge entwickelt, vielmehr nur als hydrostatischer Apparat thätig ist. In Uebereinstimmung damit finden wir den Umstand, daß die Nase bei den Fischen durch zwei blinde Gruben vorn auf der Schnautze gebildet wird, welche niemals den Gaumen durchbohren und in die Rachenhöhle münden. Dagegen sind die beiden Nasenhöhlen bei den übrigen fünf Klassen der Paarnasen zu Luftwegen umgebildet, welche den Gaumen durchbohren, und so den Lungen Luft zuführen. Die echten Fische (nach Ausschluß der Dipneusten) sind demnach die einzigen Paarnasen, welche ausschließlich durch Kiemen, und niemals durch Lungen athmen. Sie leben dem entsprechend alle im Wasser und ihre beiden Beinpaare haben die ursprüngliche Form von rudernden Flossen beibehalten.
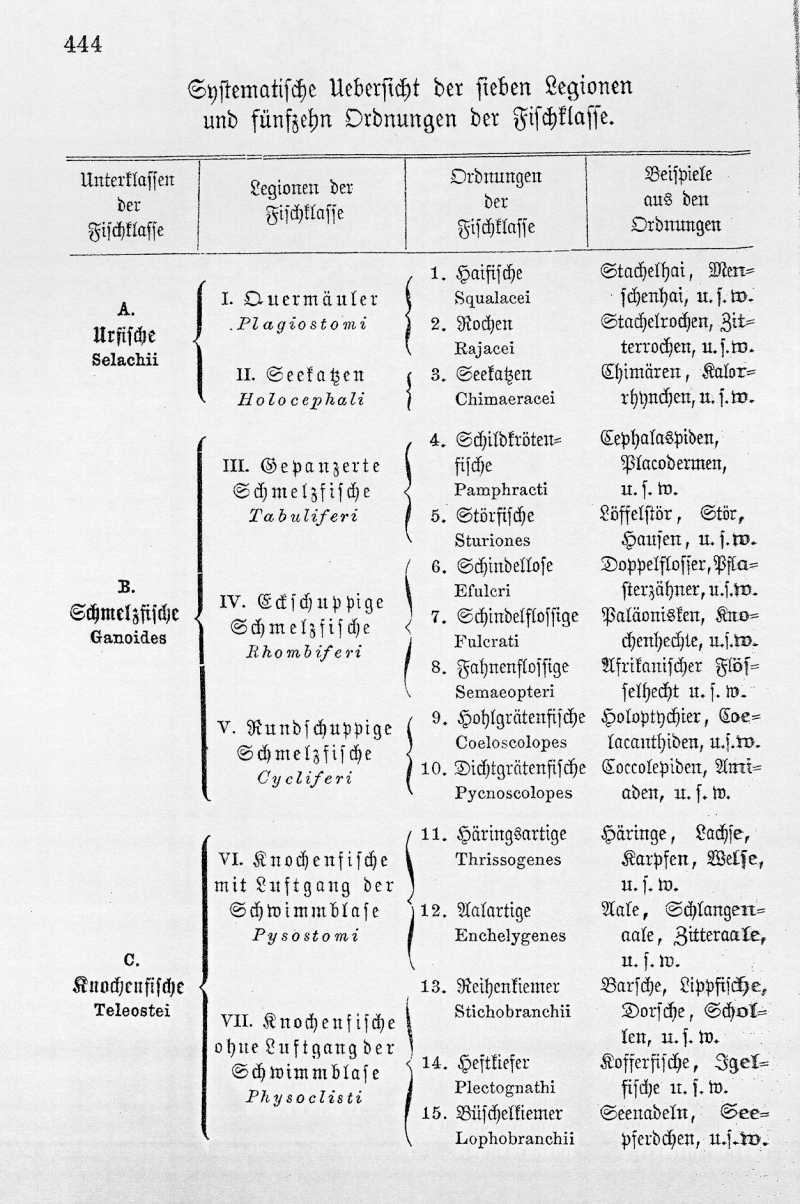
Die echten Fische werden in drei verschiedene Unterklassen eingetheilt, in die Urfische, Schmelzfische und Knochenfische. Die älteste von diesen, welche die ursprüngliche Form am getreuesten bewahrt hat, ist diejenige der Urfische (Selachii). Davon leben heutzutage noch die Haifische (Squali) und Rochen (Rajae), welche man als Quermäuler (Plagiostomi) zusammenfaßt, sowie die seltsame Fischform der abenteuerlich gestalteten Seekatzen oder Chimären (Holocephali oder Chimaeracei). Aber diese Urfische der Gegenwart, welche in allen Meeren vorkommen, sind nur schwache Reste von der gestaltenreichen und herrschenden Thiergruppe, welche die Selachier in früheren Zeiten der Erdgeschichte, und namentlich während der paläolithischen Zeit bildeten. Leider besitzen alle Urfische ein knorpeliges, niemals vollständig verknöchertes Skelet, welches der Versteinerung nur wenig oder gar nicht fähig ist. Die einzigen harten Körpertheile, welche in fossilem Zustande sich erhalten konnten, sind die Zähne und die Flossenstacheln. Diese finden sich aber in solcher Menge, Formenmannichfaltigkeit und Größe in den älteren Formationen vor, daß wir daraus mit Sicherheit auf eine höchst beträchtliche Entwickelung derselben {Tabelle Seite 444} in jener altersgrauen Vorzeit schließen können. Sie finden sich sogar schon in den silurischen Schichten, welche von anderen Wirbelthieren nur schwache Reste von Schmelzfischen (und diese erst in den jüngsten Schichten, im oberen Silur) einschließen.
Von den drei Ordnungen der Urfische sind die bei weitem wichtigsten und interessantesten die Haifische, welche wahrscheinlich unter allen lebenden Paarnasen der ursprünglichen Stammform der ganzen Gruppe, den Proselachiern, am nächsten stehen. Aus Paarnasen, welche von echten Haifischen vermuthlich nur wenig verschieden waren, haben sich als drei divergente Linien einerseits die Schmelzfische, andrerseits die Lurchfische, und drittens, als wenig veränderte Stammlinie, die übrigen Selachier entwickelt.
Die Schmelzfische (Ganoides) stehen in anatomischer Beziehung vollständig in der Mitte zwischen den Urfischen einerseits und den Knochenfischen andrerseits. In vielen Merkmalen stimmen sie mit jenen, in vielen anderen mit diesen überein. Wir ziehen daraus den Schluß, daß sie auch genealogisch den Uebergang von den Urfischen zu den Knochenfischen vermittelten. In noch höherem Maaße, als die Urfische, sind auch die Ganoiden heutzutage größtentheils ausgestorben, wogegen sie während der ganzen paläolithischen und mesolithischen Zeit in großer Mannichfaltigkeit und Masse entwickelt waren. Nach der verschiedenen Form der äußeren Hautbedeckung theilt man die Schmelzfische in drei Legionen: Gapanzerte, Eckschuppige und Rundschuppige.
Die gepanzerten Schmelzfische (Tabuliferi) sind die ältesten und schließen sich unmittelbar an die Selachier an, aus denen sie entsprungen sind. Fossile Reste von ihnen finden sich, obwohl selten, bereits im oberen Silur vor (Pteraspis ludensis aus den Ludlowschichten). Riesige, gegen 30 Fuß lange Arten derselben, mit mächtigen Knochenplatten gepanzert, finden sich namentlich im devonischen System. Heute aber lebt von dieser Legion nur noch die kleine Ordnung der Störfische (Sturiones), nämlich die Löffelstöre (Spatularides), und die Störe (Accipenserides), zu denen u. A. der Hausen gehört, welcher uns den Fischleim oder die Hausenblase liefert, der Stör und Störlett, deren Eier wir als Caviar verzehren, u. s. w.
Aus den gepanzerten Schmelzfischen haben sich wahrscheinlich als zwei divergente Zweige die eckschuppigen und die rundschuppigen entwickelt. Die eckschuppigen Schmelzfische (Rhombiferi), welche man durch ihre viereckigen oder rhombischen Schuppen auf den ersten Blick von allen anderen Fischen unterscheiden kann, sind heutzutage nur noch durch wenige Ueberbleibsel vertreten, nämlich durch den Flösselhecht (Polypterus) in afrikanischen Flüssen (vorzüglich im Nil), und durch den Knochenhecht (Lepidosteus) in amerikanischen Flüssen. Aber während der paläolithischen und der ersten Hälfte der mesolithischen Zeit bildete diese Legion die Hauptmasse der Fische. Weniger formenreich war die dritte Legion, die rundschuppigen Schmelzfische (Cycliferi), welche vorzugsweise während der Devonzeit und Steinkohlenzeit lebten. Jedoch war diese Legion, von der heute nur noch der Kahlhecht (Amia) in nordamerikanischen Flüssen übrig ist, insofern viel wichtiger, als sich aus ihnen die dritte Unterklasse der Fische, die Knochenfische, entwickelten.
Die Knochenfische (Teleostei) bilden in der Gegenwart die Hauptmasse der Fischklasse. Es gehören dahin die allermeisten Seefische, und alle unsere Süßwasserfische, mit Ausnahmen der eben erwähnten Schmelzfische. Wie zahlreiche Versteinerungen deutlich beweisen, ist diese Klasse erst um die Mitte des mesolithischen Zeitalters aus den Schmelzfischen, und zwar aus den rundschuppigen oder Cycliferen entstanden. Die Thrissopiden der Jurazeit (Thrissops, Leptolepis, Tharsis), welche unseren heutigen Häringen am nächsten stehen, sind wahrscheinlich die ältesten von allen Knochenfischen, und unmittelbar aus den rundschuppigen Schmelzfischen, welche der heutigen Amia nahe standen, hervorgegangen.
Bei den älteren Knochenfischen, den Physostomen war, ebenso wie bei den Ganoiden, die Schwimmblase noch zeitlebens durch einen bleibenden Luftgang (eine Art Luftröhre) mit dem Schlunde in Verbindung. Das ist auch heute noch bei den zu dieser Gruppe gehörigen Häringen, Lachsen, Karpfen, Welsen, Aalen u. s. w. der Fall. Während der Kreidezeit trat aber bei einigen Physostomen eine Verwachsung, ein Verschluß jenes Luftgangs ein, und dadurch wurde die Schwimmblase völlig von dem Schlunde abgeschnürt. So entstand die zweite Legion der Knochenfische, die der Physoklisten, welche erst während der Tertiärzeit ihre eigentliche Ausbildung erreichte, und bald an Mannichfaltigkeit bei weitem die Physostomen übertraf. Es gehören hierher die meisten Seefische der Gegenwart, namentlich die umfangreichen Familien der Dorsche, Schollen, Thunfische, Lippfische, Umberfische u. s. w., ferner die Heftkiefer (Kofferfische und Igelfische) und die Büschelkiemer (Seenadeln und Seepferdchen). Dagegen sind unter unseren Flußfischen nur wenige Physoklisten, z. B. der Barsch und der Stichling; die große Mehrzahl der Flußfische sind Physostomen.
Zwischen den echten Fischen und den Amphibien mitten inne steht die merkwürdige Klasse der Lurchfische oder Molchfische (Dipneusta oder Protopteri). Davon leben heute nur noch wenige Repräsentanten, nämlich der amerikanische Molchfisch (Lepidosiren paradoxa) im Gebiete des Amazonenstroms, und der afrikanische Molchfisch (Protopterus annectens) in verschiedenen Gegenden Afrikas. Während der trockenen Jahreszeit, im Sommer, vergraben sich diese seltsamen Thiere in den eintrocknenden Schlamm, in ein Nest von Blättern, und athmen dann Luft durch Lungen, wie die Amphibien.
Während der nassen Jahreszeit aber, im Winter, leben sie in Flüssen und Sümpfen, und athmen Wasser durch die Kiemen, gleich den Fischen. Aeußerlich gleichen sie aalförmigen Fischen, und sind wie diese mit Schuppen bedeckt; auch in manchen Eigenthümlichkeiten ihres inneren Baues, des Skelets, der Extremitäten etc. gleichen sie mehr den Fischen, als den Amphibien. In anderen Merkmalen dagegen stimmen sie mehr mit den letzteren überein, vor allen in der Bildung der Lungen, der Nase und des Herzens. Aus diesen Gründen herrscht unter den Zoologen ein ewiger Streit darüber, ob die Lurchfische eigentlich Fische oder Amphibien seien. Ebenso ausgezeichnete Zoologen haben sich für die eine, wie für die andere Ansicht ausgesprochen. In der That sind sie wegen der vollständigen Mischung des Charakters weder das eine noch das andere, und werden wohl am richtigsten als eine besondere Wirbelthierklasse ausgefaßt, welche den Uebergang zwischen jenen beiden Klassen vermittelt.
Die heute noch lebenden Dipneusten sind wahrscheinlich die letzten überlebenden Reste einer vormals formenreichen Gruppe, welche aber wegen Mangels fester Skelettheile keine versteinerten Spuren hinterlassen konnte. Sie verhalten sich in diser Beziehung ganz ähnlich den Monorrhinen und den Leptocardiern, mit denen sie gewöhnlich zu den Fischen gerechnet werden. Wahrscheinlich sind ausgestorbene Dipneusten der paläolithischen Periode, welche sich entweder in antedevonischer oder in devonischer oder in antocarbonischer Zeit aus Urfischen entwickelt hatten, die Stammformen der Amphibien, und somit auch aller höheren Wirbelthiere. Mindestens werden die unbekannten Uebergangsformen von den Urfischen zu den Amphibien, welche wir als Stammgruppe der letztren zu betrachten haben, den Dipneusten wohl sehr ähnlich gewesen sein.
Die Lurche (Amphibia) sind jedenfalls von den Urfischen oder Selachiern abzuleiten, entweder direct oder durch Vermittlung der Lurchfische. Wir theilen diese Klasse in zwei Unterklassen ein, in die Panzerlurche und Nacktlurche, von denen die ersteren durch die Bedeckung des Körpers mit Knochentafeln oder Schuppen ausgezeichnet sind.
Die älteren von diesen sind die Panzerlurche (Phractamphibia), die ältesten landbewohnenden Wirbelthiere, von denen uns fossile Reste erhalten sind. Wohlerhaltene Versteinerungen derselben finden sich schon in der Steinkohle vor, nämlich die den Fischen noch am nächsten stehenden Schmelzköpfe (Ganocephala), der Archegosaurus von Saabrücken, und das Dendrerpeton aus Nordamerika. Auf diese folgen dann später die riesigen Wickelzähner (Labyrinthodontia), schon im permischen System durch Zygosaurus, später aber vorzüglich in der Trias durch Mastodonsaurus, Trematosaurus, Kapitosaurus u. s. w. vertreten. Diese furchtbaren Raubthiere scheinen in der Körperform zwischen den Krokodilen, Salamandern und Fröschen in der Mitte gestanden zu haben, waren aber den beiden letzteren mehr durch ihren inneren Bau verwandt, während sie durch die feste Panzerbedeckung mit starken Knochentafeln den ersteren glichen. Schon gegen Ende der Triaszeit scheine diese gepanzerten Riesenlurche ausgestorben zu sein. Aus der ganzen folgenden Zeit kennen wir keine Versteinerungen von Panzerlurchen. Das diese Unterklasse jedoch währenddessen noch lebte und niemals ganz ausstarb, beweisen die heute noch lebenden Blindwühlen oder Caecilien (Peromela), kleine beschuppte Phraktamphibien von der Form und Lebensweise des Regenwurms.
Die zweite Unterklasse der Amphibien, die Nacktlurche (Lissamphibia), entstanden wahrscheinlich schon während der primären oder secundären Zeit, obgleich wir fossile Reste derselben erst aus der Tertiärzeit kennen. Sie unterscheiden sich von den Panzerlurchen durch ihre nackte, glatte, schlüpfrige Haut, welche jeder Schuppen- oder Panzerbedeckung entbehrt. Sie entwickelten sich vermuthlich entweder aus einem Zweige der Phraktamphibien oder aus gemeinsamer Wurzel mit diesen. Die drei Ordnungen von Nacktlurchen, welche noch jetzt leben, die Kiemenlurche, Schwanzlurche und Forschlurche, wiederholen uns noch heutzutage in ihrer individuellen Entwickelung sehr deutlich den historischen Entwickelungsgang der ganzen Unterklasse.
Die ältesten Formen sind die Kiemenlurche (Sozobranchia), welche zeitlebens auf der ursprünglichen Stammform der Nacktlurche stehen bleiben und einen langen Schwanz nebst wasserathmenden Kiemen beibehalten. Sie stehen am nächsten den Dipneusten, von denen sie sich aber schon äußerlich durch den Mangel des Schuppenkleides unterscheiden. Die meisten Kiemenlurche leben in Nordamerika, unter anderen der früher erwähnte Axolotl oder Siredon (vergl. oben S. 192). In Europa ist diese Ordnung nur durch eine Form vertreten, durch den berühmten Olm (Proteus anguineus), welcher die Adelsberger Grotte und andere Höhlen Krains bewohnt, und durch den Aufenthalt im Dunkeln rudimentäre Augen bekommen hat, die nicht mehr sehen können (S. oben S. 11). Aus den Kiemenlurchen hat sich durch Verlust der äußeren Kiemen die Ordnung der Schwanzlurche (Sozura) entwickelt, zu welcher unser schwarzer, gelbgefleckter Landsalamander (Salamandra maculata) und unsere flinken Wassermolche (Triton) gehören. Manche von ihnen, z. B. der berühmte Riesenmolch von Japan (Cryptobranchus japonicus) haben noch die Kiemenspalte beibehalten, trotzdem sie die Kiemen selbst verloren haben. Alle aber behalten den Schwanz zeitlebens. Bisweilen conserviren die Tritonen auch die Kiemen und bleiben so ganz auf der Stufe der Kiemenlurche stehen, wenn man sie nämlich zwingt, beständig im Wasser zu bleiben (Vergl. oben S. 192).
Die dritte Ordnung, die Schwanzlosen oder Froschlurche (Anura), verlieren bei der Metamorphose nicht nur die Kiemen, durch welche sie in früher Jugend (als sogenannte "Kaulquappen") Wasser athmen, sondern auch den Schwanz, mit dem sie umherschwimmen. Sie durchlaufen also während ihrer Ontogenie den Entwickelungsgang der ganzen Unterklaslse, indem sie zuerst Kiemenlurche, später Schwanzlurche, und zuletzt Froschlurche sind. Offenbar ergiebt sich daraus, daß die Froschlurche sich erst später aus den Schwanzlurchen, wie diese selbst aus den ursprünglich allein vorhandenen Kiemenlurchen entwickelt haben.
Indem wir nun von den Amphibien zu der nächsten Wirbelthierklasse, den Reptilien übergehen, bemerken wir eine sehr bedeutende Vervollkommnung in der stufenweise fortschreitenden Organisation der Wirbelthiere. Alle bisher betrachteten Paarnasen oder Amphirrhinen, nämlich die drei nahe verwandten Klassen der Fische, Lurchfische und Lurche, stimmen in einer Anzahl von wichtigen Charakteren überein, durch welche sie sich von den drei noch übrigen Wirbelthierklassen, den Reptilien, Vögeln und Säugethieren, sehr wesentlich unterscheiden. Bei diesen letzteren bildet sich während der embryonalen Entwickelung rings um den Embryo eine von seinem Nabel auswachsende besondere zarte Hülle, die Fruchthaut oder das Amnion, welche mit dem Fruchtwasser oder Amnionwasser gefüllt ist, und in diesem den Embryo oder Keim blasenförmig umschließt. Wegen dieser sehr wichtigen und charakteristischen Bildung können wir jene drei höchst entwickelten Wirbelthierklassen als Amnionthiere (Amniota) zusammenfassen. Die drei soeben betrachteten Klassen der Paarnasen dagegen, denen das Amnion, ebenso wie allen niederen Wirbelthieren (Unpaarnasen und Schädellosen), fehlt, können wir jenen als Amniolose (Anamnia) entgegensetzen.
Die Bildung der Fruchthaut oder des Amnion, durch welche sich die Reptilien, Vögel und Säugethiere von allen anderen Wirbelthieren unterscheiden, ist offenbar ein höchst wichtiger Vorgang in der Ontogenie und der ihr entsprechenden Phylogenie der Wirbelthiere. Er fällt zusammen mit einer Reihe von anderen Vorgängen, welche wesentlich die höhere Entwickelung der Amnionthiere bestimmten. Dahin gehört vor allen der gänzliche Verlust der Kiemen, dessenwegen man schon früher die Amnioten als Kiemenlose (Ebranchiata) allen übrigen Wirbelthieren als Kiemenathmenden (Branchiata) entgegengesetzt hatte.
Bei allen bisher betrachteten Wirbelthieren fanden sich athmende Kiemen entweder zeitlebens, oder doch wenigstens, wie bei Fröschen und Molchen, in früher Jugend. Bei den Reptilien, Vögeln und Säugethieren dagegen kommen zu keiner Zeit des Lebens wirklich athmende Kiemen vor, und die auch hier vorhandenen Kiemenbogen gestalten sich im Laufe der Ontogenie zu ganz anderen Gebilden, zu Theilen des Kieferapparats und des Gehörgangs (Vergl. oben S. 251). Alle Amnionthiere besitzen im Gehörgang eine sogenannte "Schnecke" und ein dieser entsprechendes "rundes Fenster". Diese Theile fehlen dagegen bei den Amnionlosen.
Bie diesen letzteren liegt der Schädel des Embryo in der gradlinigen Fortsetzung der Wirbelsäule. Bei den Amnionthieren dagegen erscheint die Schädelbasis von der Bauchseite her eingeknickt, so daß der Kopf auf die Brust herabsinkt (S. 240 c, d, Fig. A-E). Auch entwickeln sich erst bei den Amnioten die Thränenorgane im Auge, welche den Anamnien noch fehlen. Wann fand nun im Laufe der organischen Erdgeschichte dieser wichtige Vorgang statt? Wann entwickelte sich aus einem Zweige der Amnionlosen (und zwar jedenfalls aus einem Zweige der Amphibien) der gemeinsame Stammvater aller Amnionthiere?
Auf diese Frage geben uns die versteinerten Wirbelthierreste zwar keine ganz bestimmte, aber doch eine annähernde Antwort. Mit Ausnahme nämlich von zwei im permischen Systeme gefundenen eidechsenähnlichen Thieren (dem Proterosaurus und Rhopalodon) gehören alle übrigen versteinerten Reste, welche wir bis jetzt von Amnionthieren kennen, der Secundärzeit, Tertiärzeit und Quartärzeit an. Von jenen beiden Wirbelthieren aber ist es noch zweifelhaft, ob sie schon wirkliche Reptilien und nicht vielleicht salamanderähnliche Amphibien sind. Wir kennen von ihnen allein das Skelet und dies nicht einmal vollständig. Im Ganzen gleicht das Skelet allerdings mehr den Reptilien als den Amphibien, in manchen Einzelheiten mehr den Amphibien.
Da wir nun von den entscheidenden Merkmalen der Weichtheile gar Nichts wissen, ist es sehr wohl möglich, daß der Proterosaurus und der Rhopalodon noch amnionlose Thiere waren, welche den Amphibien näher als den Reptilien standen, vielleicht aber zu den Uebergangsformen zwischen beiden Klassen gehörten. Da aber andrerseits unzweifelhafte Amnionthiere bereits in der Trias versteinert vorgefunden werden, so ist es wahrscheinlich, daß die Hauptklasse der Amnioten sich erst in der Antetriaszeit, im Beginn des mesolithischen Zeitalters, entwickelte. Wie wir schon früher sahen, ist offenbar gerade dieser Zeitraum einer der wichtigsten Wendepunkte in der organischen Erdgeschichte. An die Stelle der paläolitischen Farnwälder traten damals die Nadelwälder der Trias. In vielen Abtheilungen der wirbellosen Thiere traten wichtige Umgestaltungen ein: Aus den getäfelten Seelilien (Phatnocrina) entwickelten sich die gegliederten (Colocrina). die Autechiniden oder die Seeigel mit zwanzig Plattenreihen traten an die Stelle der paläolithischen Palechiniden, der Seeigel mit mehr als zwanzig Plattenreihen. Die Cystiden, Blastoideen, Trilobiten und andere charakteristische wirbellose Thiergruppen der Primärzeit waren so eben ausgestorben. Kein Wunder, wenn die umgestalteten Anpassungsverhältnisse der Antetriaszeit auch auf den Wirbelthierstamm mächtig einwirkten, und die Entstehung der Amnionthiere veranlaßten.
Wenn man dagegen die beiden eidechsen- oder salamanderähnlichen Thiere der Permzeit, den Proterosaurus und den Rhopalodon, als echte Reptilien, mithin als die ältesten Amnioten betrachtet, so würde die Entstehung dieser Hauptklasse bereits um eine oder zwei Perioden früher, gegen das Ende der Primärzeit, fallen, in die vermischte oder antepermische Periode. Alle übrigen Reptilienreste aber, welche man früher im permischen, im Steinkohlensystem oder gar im devonischen Systeme gefunden zu haben glaubte, haben sich entweder nicht als Reptilienreste, oder als viel jüngeren Alters (meistens der Trias angehörig) herausgestellt.
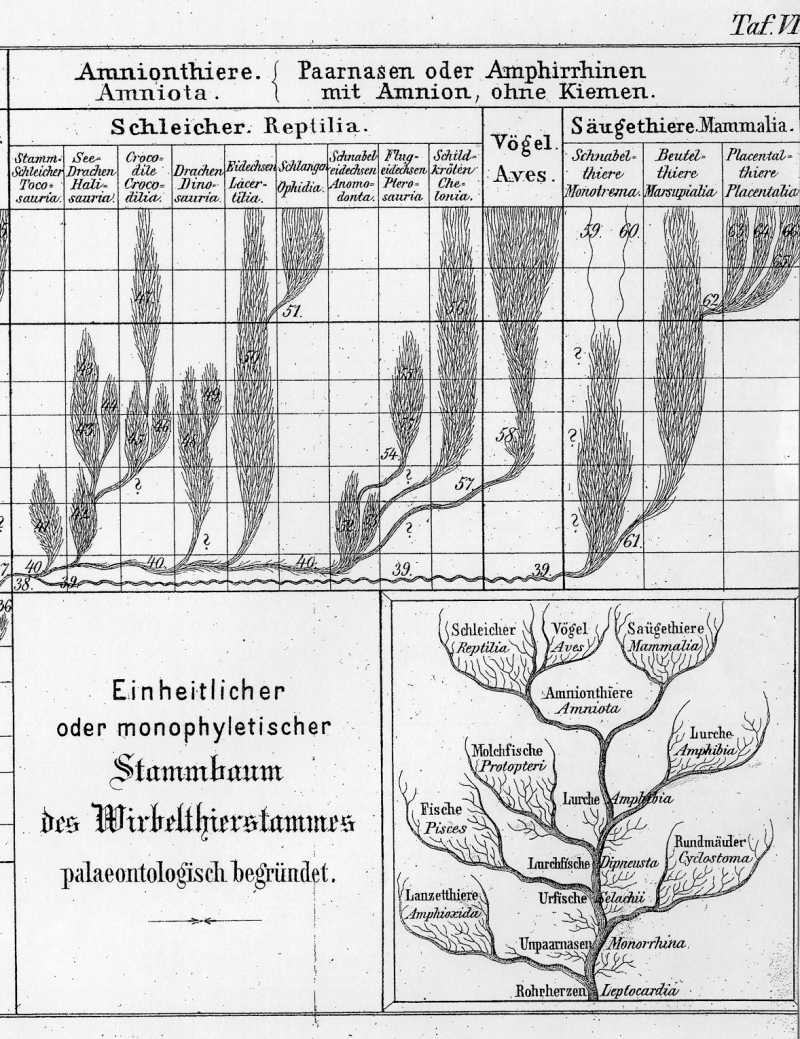
Die gemeinsame hypothetische Stammform aller Amnionthiere, welche wir als Protamnion bezeichnen können, und welche möglicherweise dem Proterosaurus sehr nahe verwandt war, stand vermuthlich im Ganzen hinsichtlich ihrer Körperbildung in der Mitte zwischen den Salamandern und Eidechsen. Ihre Nachkommenschaft spaltete sich schon frühzeitig in zwei verschiedene Linien (Taf. VI, 39, 40), von denen die eine die gemeinsame Stammform der Reptilien und Vögel, die andere die Stammform der Säugethiere wurde.
Die Schleicher (Reptilia oder Pholidota, auch Sauria im weitesten Sinne genannt) bleiben von allen drei Klassen der Amnionthiere auf der tiefsten Bildungsstufe stehen und entfernen sich am wenigsten von ihren Stammvätern, den Amphibien. Daher wurden sie früher allgemein zu diesen gerechnet, obwohl sie in ihrer ganzen Organisation viel näher den Vögeln als den Amphibien verwandt sind. Gegenwärtig leben von den Reptilien nur noch vier Ordnungen, nämlich die Eidechsen, Schlangen, Krokodile und Schildkröten. Diese bilden aber nur noch einen schwachen Rest von der ungemein mannichfaltig und bedeutend entwickelten Reptilienschaar, welche während der mesolithischen oder Secundärzeit lebte und damals alle anderen Wirbelthierklassen beherrschte. Die ausnehmende Entwickelung der Reptilien während der Secundärzeit ist so charakteristisch, daß wir diese darnach eben so gut, wie nach den Gymnospermen benennen könnten (S. 306). Von den dreißig Unterordnungen, welche die nachstehende Tabelle Ihnen vorführt, gehört die Hälfte, und von den neun Ordnungen gehören fünf ausschließlich der Secundarzeit an .Auch von den fünf Unterklassen, auf welche wir jene vertheilen können, sind zwei (I und IV) gänzlich, und zwei andere (II und V) größtentheils ausgestorben. Diese mesolithischen Gruppen sind durch ein {Kreuz} bezeichnet.
In der ersten Unterklasse, den Stammreptilien oder Stammschleichern (Tocosauria), fassen wir die ausgestorbenen Fachzähner (Thecodontia) der Triaszeit mit denjenigen Reptilien zusammen, welche wir als die gemeinsame Stammform der ganzen Klasse betrachten können. Zu diesen letzteren, welche wir als Urschleicher (Proreptilia) bezeichnen können, gehört möglicherweise der Proterosaurus des permischen Systems. Die vier übrigen Unterklassen sind wahrscheinlich als vier divergente Zweige aufzufassen, welche sich aus jener gemeinsamen Stammform nach verschiedenen Richtungen hin entwickelt haben. Die Thecodonten der Trias, die einzigen sicher bekannten fossilen Reste von Tocosauriern, waren Eidechsen, welche den heute noch lebenden Monitoren oder Warneidechsen (Monitor, Varanus) ziemlich ähnlich gewesen zu sein scheinen.
Die zweite Unterklasse, die Schwimmschleicher (Hydrosauria), lebte ganz oder größtentheils im Wasser. Sie spaltet sich in die beiden Ordnungen der Seedrachen und der Krokodile. Die riesigen, bis 40 Fuß langen Seedrachen (Halisauria) lebten bloß während der Secundärzeit, und zwar finden sich die versteinerten Reste der Simosaurier bloß in der Trias, diejenigen der Plesiosauier und Ichthyosaurier bloß im Jura und in der Kreide. Vermuthlich entwickelten sich also die letzteren aus den ersteren während der Antejurazeit. Um diese Zeit entstanden wahrscheinlich auch die Krokodile (Crocodilia), von denen die Teleosaurier und Steneosaurier bloß im Jura, die jetzt allein noch lebenden Alligatoren aber in den Kreide und Tertiärschichten versteinert gefunden werden. Die wenigen Krokodile der Gegenwart sind nur ein dürftiger Rest von der furchtbaren Raubthierschaar, welche die Gewässer der mesolithischen Zeit bevölkerte.
Von allen Reptiliengruppen hat sich bis auf unsere Zeit am besten die dritte Unterklasse conservirt, die Schuppenschleicher (Lepidosauria). Es gehören dahin die beiden nächstverwandten Ordnungen { Tabelle Seite 455 } der echten Eidechsen (Lacertilia) und der Schlangen (Ophidia), von denen jede in fünf verschiedene Unterordnungen zerfällt. Unter den echten Eidechsen stehen die Monitoren oder Warneidechsen (Monitor, Varanus) den ursprünglichen Stammformen der ganzen Klasse am nächsten. Die Schlangen, die jüngste von allen neun Reptilienordnungen, scheinen sich erst während der Anteeocenzeit, im Beginn der Tertiärzeit, aus einem Zweige der Eidechsen entwickelt zu haben. Wenigstens kennt man versteinerte Schlangen bis jetzt bloß aus tertiären Schichten. Gänzlich ausgestorben, ohne Nachkommen zu hinterlassen, ist die vierte Unterklasse, diejenige der Drachen oder Lindwürmer (Dinosaurier oder Pachypoda).

Diese kolossalen Reptilien, welche eine Länge von mehr als 50 Fuß erreichten, sind die größten Landbewohner, welche jemals unser Erdball getragen hat. Sie lebten ausschließlich in der Secundärzeit. Die meisten Reste derselben finden sich in der unteren Kreide, namentlich in der Wälderformation Englands. Die Mehrzahl waren furchtbare Raubthiere (Megalosaurus von 20-30, Pelorosaurus von 40-50 Fuß Länge). Iguanodon jedoch und einige andere lebten von Pflanzennahrung und spielten in den Wäldern der Kreidezeit wahrscheinlich eine ähnliche Rolle, wie die ebenso schwerfälligen, aber kleineren Elephanten, Flußpferde und Nashörner der Gegenwart.
In einer fünften und letzten Unterklasse, Schnabelschleicher (Rhamphosauria), vereinigen wir alle diejenigen Reptilien, bei denen die Kiefer sich mehr oder weniger deutlich zu einem Vogelschnabel umbilden. Die Zähne gehen dabei ganz oder theilweise verloren, oder werden eigenthümlich umgebildet. Als gemeinsame Stammgruppe derselben, die sich aus einem oder mehreren Aesten der Tocosauerier entwickelte, können wir die Schnabeleidechsen (Anomodonta) der älteren Secundärzeit betrachten, von denen sich viele merkwürdige Reste in der Trias und im Jura finden. Aus diesen haben sich vielleicht als drei divergente Zweige die Flugschleicher, Schildkröten und Vögel entwickelt. Die merkwürdigen Flugschleicher (Pterosaurier), bei denen der außerordentlich verlängerte fünfte Finger der Hand als Stütze einer gewaltigen Flughaut diente, flogen in der Secundärzeit wahrscheinlich in ähnlicher Weise umher, wie jetzt die Fledermäuse.
Die kleinsten Flugeidechsen hatten die Größe eines Sperlings. Die größten aber, mit einer Klafterweite der Flügel von mehr als 16 Fuß, übertrafen die größten jetzt lebenden fliegenden Vögel (Condor und Albatros) an Umfang. Ihre versteinerten Reste, die langschwänzigen Rhamphorhynchen und die kurzschwänzigen Pterodactylen, finden sich zahlreich versteinert in allen Schichten der Jura- und Kreidezeit, aber nur in diesen vor. Dagegen finden wir versteinerte Schildkröten (Chelonia) vom Jura an in allen secundären, tertiären und quartären Schichten versteinert vor. Doch sind auch die Schildkröten der Gegenwart, gleich den meisten anderen Reptiliengruppen, nur schwache Ueberreste ihres früheren Glanzes. In den Tertiärschichten des Himalaya fand sich unter anderen eine versteinerte Schildkröte, die gegen 20 Fuß lang und 6 Fuß hoch war.
Die Klasse der Vögel (Aves) ist, wie schon bemerkt, durch ihren inneren Bau und durch ihre embryonale Entwickelung den Reptilien so nahe verwandt, daß sie zweifelsohne aus einem Zweige dieser Klasse ihren wirklichen Ursprung genommen hat. Wie Ihnen allein schon ein Blick auf Fig. C-F, S. 242 zeigt, sind die Embryonen der Vögel zu einer Zeit, in der sie bereits sehr wesentlich von den Embryonen der Säugethiere verschieden erscheinen, von denen der Schildkröten und anderer Reptilien noch kaum zu unterscheiden.
Die Dotterfurchung ist bei den Vögeln und Reptilien partiell, bei den Säugethieren dagegen nicht. Die Haare der Säugethiere entwickeln sich in geschlossenen Bälgen der Haut, die Federn der Vögel dagegen, eben so wie die Schuppen der Reptilien, auf Höckern der Haut. Der Unterkiefer der letzteren ist viel verwickelter zusammengesetzt, als derjenige der Säugethiere. Auch fehlt diesen letzteren das Quadratbein der ersteren. Während bei den Säugethieren (wie bei den Amphibien) die Verbindung zwischen dem Schädel und dem ersten Halswirbel durch zwei Gelenkhöcker oder Condylen geschieht, sind diese dagegen bei den Vögeln und Reptilien zu einem einzigen verschmolzen. Man kann die beiden letzteren Klassen daher mit vollem Rechte in einer Gruppe als Monocondyla zusammenfassen und dieser die Säugethiere als Dicondyla gegenüber setzen.
Die Abzweigung der Vögel von den Reptilien fand jedenfalls erst während der mesolithischen Zeit, und zwar wahrscheinlich während der Triaszeit oder Antejurazeit statt. Die ältesten fossilen Vogelreste sind im oberen Jura gefunden worden (Archaepteryx). Aber schon in der Triaszeit lebten verschiedene Saurier (Anomodonten), die in mehrfacher Hinsicht den Uebergang von den Tocosauriern zu den Stammvätern der Vögel, den hypothetischen Tocornithen, zu bilden scheinen. Wahrscheinlich waren diese Tocornithen von anderen Schnabeleidechsem im Systeme kaum zu trennen, und namentlich dem känguruhartigen Compsognathus aus dem Jura von Solnhofen nächst verwandt. Huxley stellt diesen letzteren zu den Dinosauriern, und glaubt, daß diese die nächsten Verwandten der Tocornithen seien.
Die große Mehrzahl der Vögel erscheint, trotz aller Mannichfaltigkeit in der Färbung des schönen Federkleides und in der Bildung des Schnabels und der Füße, höchst einförmig organisirt, in ähnlicher Weise, wie die Insectenklasse. Den äußeren Existenzbedingungen hat sich die Vogelform auf das Vielfältigste angepaßt, ohne dabei irgend wesentlich von dem streng erblichen Typus der charakteristischen inneren Bildung abzuweichen. Nur zwei kleine Gruppen, einerseits die fiederschwänzigen Vögel (Saururae), andrerseits die straußartigen (Ratidae), weichen erheblich von dem gewöhnlichen Vogeltypus, dem der kielbrüstigen (Carinatae) ab, und demnach kann man die ganze Klasse in drei Unterklassen eintheilen.
Die erste Unterklasse, die reptilienschwänzigen oder fiederschwänzigen Vögel (Saururae) sind bis jetzt bloß durch einen einzigen und dazu unvollständigen fossilen Abdruck bekannt, welcher aber als die älteste und dabei sehr eigenthümliche Vogelversteinerung eine hohe Bedeutung beansprucht. Das ist der Urgreif oder die Archaeopteryx lithographica, welche bis jetzt erst in einem Exemplar in dem lithographischen Schiefer von Solenhofen, im oberen Jura von Baiern, gefunden wurde. Dieser merkwürdige Vogel scheint im Ganzen Größe und Wuchs eines starken Raben gehabt zu haben, namentlich was die wohl erhaltenen Beine betrifft; Kopf und Brust fehlen leider. Die Flügelbildung weicht schon etwas von derjenigen der anderen Vögel ab, noch viel mehr aber der Schwanz. Bei allen übrigen Vögeln ist der Schwanz sehr kurz, aus wenigen kurzen Wirbeln zusammengesetzt. Die letzten derselben sind zu einer dünnen senkrecht stehenden Knochenplatte verwachsen, an welcher sich die Steuerfedern des Schwanzes fächerförmig ansetzen.
Die Archäopteryx dagegen hat einen langen Schwanz, wie die Eidechsen, aus zahlreichen (20) langen und dünnen Wirbeln zusammengesetzt, und an jedem Wirbel sitzen zweizeilig ein paar starke Steuerfedern, so daß der ganze Schwanz regelmäßig gefiedert erscheint. Dieselbe Bildung der Schwanzwirbelsäule zeigt sich bei den Embryonen der übrigen Vögel vorübergehend, so daß offenbar der Schwanz der Archäopteryx die ursprüngliche, von den Reptilien ererbte Form des Vogelschwanzes darstellt. Wahrscheinlich lebten ähnliche Vögel mit Eidechsenschwanz um die mittlere Secundärzeit in großer Menge; der Zufall hat uns aber erst diesen einen Rest bis jetzt enthüllt.
Zu den fächerschwänzigen oder kielbrüstigen Vögeln (Carinatae), welche die zweite Unterklasse bilden, gehören alle jetzt lebenden Vögel, mit Ausnahme der straußartigen oder Ratiden. Sie haben sich wahrscheinlich in der zweiten Hälfte der Secundärzeit, in der Antekretazeit oder in der Kreidezeit, aus den fiederschwänzigen durch Verwachsung der hinteren Schwanzwirbel und Verkürzung des Schwanzes entwickelt. Aus der Secundärzeit kennt man von ihnen nur sehr wenige Reste, und zwar nur aus dem letzten Abschnitt derselben, aus der Kreide. Diese Reste gehören einem albatrosartigen Schwimmvogel und einem schnepfenartigen Stelzvogel an. Alle übrigen bis jetzt bekannten versteinerten Vogelreste sind in den Tertiärschichten gefunden worden, und zeigen, daß die Klasse erst in der Tertiärzeit ihre eigentliche Entwickelung und Ausbreitung erreichte.
Die straußartigen oder flaumschwänzigen Vögel (Ratitae), auch Laufvögel (Cursores) genannt, die dritte und letzte Unterklasse, ist gegenwärtig nur noch durch wenige lebende Arten vertreten, durch den zweizehigen afrikanischen Strauß, den dreizehigen amerikanischen und neuholländischen Strauß, den indischen Casuar, und den vierzehigen Kiwi oder Apteryx von Neuseeland. Auch die ausgestorbenen Riesenvögel von Madagaskar (Aepyornis) und von Neuseeland (Dinornis), welche viel größer waren als die jetzt lebenden größten Strauße, gehören zu dieser Gruppe. Wahrscheinlich sind die straußartigen Vögel durch Abgewöhnung des Fliegens, durch die damit verbundene Rückbildung der Flugmuskeln und des denselben zum Ansatz dienenden Brustbeinkammes, und durch entsprechend stärkere Ausbildung der Hinterbeine zum Laufen, aus einem Zweige der kielbrüstigen Vögel entstanden. Vielleicht sind dieselben jedoch auch, wie Huxley meint, nächste Verwandte der Dinosaurier, und der diesen nahestehenden Reptilien, namentlich des Compsognathus. In diesen Falle würden die Kielbrüstigen erst später aus den straußartigen, als der ursprünglichen Stammgruppe der Klasse entstanden sein.
Da die nähere Betrachtung der geschichtlichen und genealogischen Entwickelung der einzelnen Vogelordnungen gar kein besonderes Interesse hat, wenden wir uns nun sogleich zum Stammbaum der achten und letzten Wirbelthierklasse, der Säugethiere (Mammalia). Ohne Zweifel ist dies die bei weitem interessanteste, vollkommenste und wichtigste von allen Thierklassen. Denn in diese Klasse reiht die wissenschaftliche Zoologie auch den Menschen ein, und aus Gliedern dieser Klasse hat sich das Menschengeschlecht zunächst entwickelt. Wir müssen daher der Geschichte und dem Stammbaum der Säugethiere unsere besondere Aufmerksamkeit zuwenden. Lassen Sie uns zu diesem Zwecke wieder zunächst das System dieser Thierklasse untersuchen.
Von den älteren Naturforschern wurde die Klasse der Säugethiere mit vorzüglicher Rücksicht auf die Bildung des Gebisses und der Füße in einer Reihe von 8-16 Ordnungen eingetheilt. Auf der tiefsten Stufe dieser Reihe standen die Walfische, welche durch ihre fischähnliche Körpergestalt sich am meisten von den Menschen, der höchsten Stufe zu entfernen schienen. So unterschied Linné folgende acht Ordnungen:
1 Cete (Wale);
2. Belluae (Flußpferde und Pferde);
3. Pecora (Wiederkäuer);
4. Glires (Nagethiere und Nashorn);
5. Bestiae (Insectenfresser, Beutelthiere und verschiedene Andere;
6. Ferae (Raubthiere);
7. Bruta (Zahnarme und Elephanten);
8. Primates (Fledermäuse, Halbaffen, Affen und Menschen).
Nicht viel über diese Klassification von Linné erhob sich diejenige von Cuvier, welche für die meisten Zoologen maßgebend wurde. Cuvier unterschied folgende acht Ordnungen: 1. Cetacea (Wale); 2. Ruminantia (Wiederkäuer); 3. Pachyderma (Hufthiere nach Ausschluß der Wiederkäuer); 4. Edentata (Zahnarme); 5. Rodentia (Nagethiere); 6. Carnassia (Beutelthiere, Raubthiere, Insectenfresser und Flederthiere); 7. Quadrumana (Halbaffen und Affen); 8. Bimana (Menschen).
Den bedeutendsten Fortschritt in der Klassification der Säugethiere that schon 1816 der ausgezeichnete, bereits vorher erwähnte Anatom Blainville, welcher zuerst mit tiefem Blick die drei natürlichen Hauptgruppen oder Unterklassen der Säugethiere erkannte, und sie nach der Bildung ihrer Fortpflanzungsorgane als Ornithodelphien, Didelphien und Monodelphien unterschied. Da diese Eintheilung heutzutage mit Recht bei allen wissenschaftlichen Zoologen wegen ihrer tiefen Begründung durch die Entwickelungsgeschichte als die beste gilt, so lassen Sie uns derselben auch hier folgen.
Die erste Unterklasse bilden die Kloakenthiere oder Brustlosen, auch Gabler oder Gabelthiere genannt (Ornithodelphia oder Amasta). Sie sind heutzutage nur noch durch zwei lebende Säugthierarten vertreten, die beide auf Neuholland und das benachbarte Vandiemensland beschränkt sind: das wegen seines Vogelschnabels sehr bekannte Wasserschnabelthier (Ornithorhynchus paradoxus) und das wenige bekannte, igelähnliche Landschnabelthier (Echidna hystrix). Diese beiden seltsamen Thiere, welche man in der Ordnung der Schnabelthiere (Monotrema) zusammenfaßt, sind offenbar die letzten überlebenden Reste einer vormals formenreichen Thiergruppe, welche in der älteren Secundärzeit allein die Säugethierklasse vertrat, und aus der sich erst später, wahrscheinlich in der Jurazeit, die zweite Unterklasse, die Didelphien entwickelten. Leider sind uns von dieser ältesten Stammgruppe der Säugethiere, welche wir als Stammsäuger (Promammalia) bezeichnen wollen, bis jetzt noch keine fossilen Reste mit voller Sicherheit bekannt. Doch gehören dazu möglicherweise die ältesten bekannten von allen versteinerten Säugethierresten, nämlich der Microlestes antiquus, von dem man bis jetzt allerdings nur einige kleine Backzähne kennt. Diese sind in den obersten Schichten der Trias, im Keuper, und zwar zuerst (1847) in Deutschland (bei Degerloch unweit Stuttgart), später auch (1858) in England (bei Frome) gefunden worden.
Aehnliche Zähne sind neuerdings auch in der nordamerikanischen Trias gefunden und als Dromatherium sylvestre beschrieben. Diese merkwürdigen Zähne, aus deren charakteristischer Form man auf ein insectenfressendes Säugethier schließen kan, sind die einzigen Reste von Säugethieren, welche man bis jetzt in den älteren Tertiärschichten, in der Trias gefunden hat. Vielleicht gehörten aber außer diesen auch noch manche andere, im Jura und der Kreide gefundene Säugethierzähne, welche jetzt gewöhnlich Beutelthieren zugeschrieben werden, eigentlich Kloakenthieren an. Bei dem Mangel der charakteristischen Weichtheile läßt sich dies nicht sicher entscheiden. Jedenfalls müssen dem Auftreten der Beutelthiere zahlreiche, mit entwickeltem Gebiß und mit einer Kloake versehene Gabelthiere vorangegangen sein.
Die Bezeichnung: "Kloakenthiere" (Monotrema) im weiteren Sinne haben die Ornithodelphien wegen der Kloake erhalten, durch deren Besitz sie sich von allen übrigen Säugethieren unterscheiden, und degegen mit den Vögeln, Reptilien, Amphibien, überhaupt mit den niederen Wirbelthierarten übereinstimmen. Die Kloakenbildung besteht darin,daß der letzte Abschnitt des Darmkanals die Mündungen des Urogenitalapparats, d. h. der vereinigten Harn- und Geschlechtsorgane aufnimmt, während diese bei allen übrigen Säugethieren (Didelphien sowohl als Monodelphien) getrennt vom Mastdarm ausmünden. Jedoch ist auch bei diesen in der ersten Zeit des Embryolebens die Kloakenbildung vorhanden, und erst später (beim Menschen gegen die zwölfte Woche der Entwicklung) tritt die Trennung der beiden Mündungsöffnungen ein. "Gabelthiere" hat man die Kloakenthiere auch wohl genannt, weil die vorderen Schlüsselbeine mittelst des Brustbeines mit einander in der Mitte zu einem Knochenstück verwachsen sind, ähnlich dem bekannten "Gabelbein" der Vögel. Bei den übrigen Säugethieren bleiben die beiden Schlüsselbeine vorn völlig getrennt, und verwachsen nicht mit dem Brustbein. Ebenso sind die hinteren Schlüsselbeine oder Coracoidknochen bei den Gabelthieren viel stärker als bei den übrigen Säugethieren entwickelt und verbinden sich mit dem Brustbein.
Auch in vielen übrigen Charakteren, namentlich in der Bildung der inneren Geschlechtsorgane, des Gehörlabyrinthes und des Gehirns, schließen sich die Schnabelthiere näher den übrigen Wirbelthieren als den Säugethieren an, so daß man sie selbst als eine besondere Klasse von diesen hat trennen wollen. Jedoch gebären sie, gleich allen anderen Säugethieren, lebendige Junge, welche eine Zeitlang von der Mutter mit ihrer Milch ernährt werden. Während aber bei allen übrigen die Milch durch die Saugwarzen oder Zitzen der Milchdrüse entleert wird, fehlen diese bei den Schnabelthieren gänzlich, und die Milch tritt einfach aus einer Hautspalte hervor. Man kann sie daher auch als Brustlose oder Zitzenlose (Amasta) bezeichnen.
Die auffallende Schnabelbildung der beiden noch lebenden Schnabelthiere, welche mit Verkümmerung der Zähne verbunden ist, muß offenbar nicht als wesentliches Merkmal der ganzen Unterklasse der Kloakenthiere, sondern als ein zufälliger Anpassungscharakter angesehen werden, welche die letzten Reste der Klasse von der ausgestorbenen Hauptgruppe ebenso unterscheidet, wie die Bildung eines ähnlichen zahnlosen Rüssels manche Zahnarme (z. B. die Ameisenfresser) vor den übrigen Placentalthieren auszeichnet. Die unbekannten ausgestorbenen Stammsäugethiere oder Promammalien, die in der Triaszeit lebten, und von denen die beiden heutigen Schnabelthiere nur einen einzelnen, verkümmerten und einseitig ausgebildeten Ast darstellen, besaßen wahrscheinlich ein sehr entwickeltes Gebiß, gleich den Beutelthieren, die sich zunächst aus ihnen entwickelten.
Die Beutelthiere oder Beutler (Didelpha oder Marsupialia), die zweite von den drei Unterklassen der Säugethiere, vermittelt in jeder Hinsicht, sowohl in anatomischer und embryologischer, als in genealogischer und historischer Beziehung, den Uebergang zwischen den beiden anderen, den Kloakenthieren und Placentalthieren. Zwar leben von dieser Gruppe noch jetzt zahlreiche Vertreter, namentlich die allbekannten Kängurus, Beutelratten und Beutelhunde. Allein im Ganzen geht offenbar auch diese Unterklasse, gleich der vorhergehenden, ihrem völligen Aussterben entgegen, und die noch lebenden Glieder derselben sind die letzten überlebenden Reste einer großen und formenreichen Gruppe, welche während der älteren Tertiärzeit vorzugsweise die Säugethierklasse vertrat.
Wahrscheinlich haben sich die Beutelthiere um die Mitte der mesolithischen Zeit (während der Juraperiode?) aus einem Zweige der Kloakenthiere entwickelt, und im Beginn der Tertiärzeit ging wiederum aus den Beutelthieren die Gruppe der Placentalthiere hervor, welcher die ersteren dann bald im Kampfe um's Dasein unterlagen. Alle fossilen Reste von Säugethieren, welche wir aus der Secundärzeit kennen, gehören entweder ausschließlich Beutelthieren, oder (zum Theil vielleicht?) Kloakenthieren an. Damals scheinen Beutelthiere über die ganze Erde verbreitet zu sein. Selbst in Europa (England, Frankreich) finden wir zahlreiche Reste derselben. Dagegen sind die letzen Ausläufer der Unterklasse, welche jetzt noch leben, auf ein sehr enges Verbreitungsgebiet beschränkt, nämlich auf Neuholland, auf den australischen und einen kleinen Theil des asiatischen Archipelagus. Einige wenige Arten leben auch noch in Amerika; hingegen lebt in der Gegewart kein einziges Beutelthier mehr auf dem Festlande von Asien, Afrika und Europa.
Die Beutelthiere führen ihren Namen von der bei den meisten wohl entwickelten beutelförmigen Tasche (Marsupium), welche sich an der Bauchseite der weiblichen Thiere vorfindet, und in welcher die Mutter ihre Jungen noch eine geraume Zeit lang nach der Geburt umherträgt. Dieser Beutel wird durch zwei charakteristische Beutelknochen gestützt, welche auch den Schabelthieren zukommen, den Placentalthieren dagegen fehlen. Das junge Beutelthier wird in viel unvollkommener Gestalt geboren, als das junge Placentalthier, und erreicht erst, nachdem es einige Zeit im Beutel sich entwickelt hat, denjenigen Grad der Ausbildung, welchen das letztere schon gleich bei seiner Geburt besitzt. Bei dem Riesenkänguruh, welches Mannshöhe erreicht, ist das neugeborene Junge, welches nicht viel über fünf Wochen von der Mutter im Fruchbehälter getragen wurde, nicht mehr als zolllang, und erreicht seine wesentliche Ausbildung erst in dem Beutel der Mutter, wo es gegen neun Monate, an der Zitze der Milchdrüse festgesaugt, hängen bleibt.
Die verschiedenen Abtheilungen, welche man gewöhnlich als sogenannte Familien in der Unterklasse der Beutelthiere unterscheidet, verdienen eigentlich den Rang von selbstständigen Ordnungen, da sie sich in der mannichfaltigen Differenzirung des Gebisses und der Gliedmaßen in ähnlicher Weise, wenn auch nicht so scharf, von einander unterscheiden, wie die verschiedenen Ordnungen der Placentalthiere.
Zum Theil entsprechen sie den letzteren vollkommen. Offenbar hat die Anpassung an ähnliche Lebensverhältnisse in den beiden Unterklassen der Marsupialien und Placentalien ganz entsprechende oder analoge Umbildungen der ursprünglichen Grundform bewirkt. Man kann in dieser Hinsicht ungefähr acht Ordnungen von Beutelthieren unterscheiden, von denen die eine Hälfte die Hauptgruppe oder Legion der pflanzenfressenden, die andere Hälfte die Legion der fleischfressenden Marsupialien bildet. Von beiden Legionen finden sich (falls man nicht auch den vorher erwähnten Mikrolestes und das Dromatherium der Trias hierher ziehen will) die ältesten fossilen Reste im Jura vor, und zwar in den Schiefern von Stonesfield, bei Oxford in England. Diese Schiefer gehören der Bathformation oder dem unteren Oolith an, derjenigen Schichtengruppe, welche unmittelbar über dem Lias, der ältesten Jurabildung liegt (Vergl. S. 307).
Allerdings bestehen die Beutelthierreste, welche in den Schiefern von Stonesfield gefunden wurden, und ebenso diejenigen, welche man später in den Purbeckschichten fand, nur aus Unterkiefern (Vergl. S. 311). Allein glücklicherweise gehört gerade der Unterkiefer zu dem am meisten charakteristischen Skelettheilen der Beutelthiere. Er zeichnet sich nämlich durch einen hakenförmigen Fortsatz des nach unten und hinten gekehrten Unterkieferwinkels aus, welcher weder den Placentalthieren noch den (heute lebenden) Schnabelthieren zukömmt, und wir können aus der Anwesenheit dieses Fortsatzes an den Unterkiefern von Stonesfield schließen, daß sie Beutelthieren angehört haben.
Von den pflanzenfressenden Beutelthieren (Botanophaga) kennt man bis jetzt aus dem Jura nur zwei Versteinerungen, nämlich den Stereognathus oolithicus aus den Schiefern von Stonesfield (unterer Oolith) und den Plagiaulax Becklesii aus den mittleren Purbeckschichten (oberer Oolith). Dagegen finden sich in Neuholland riesige versteinerte Reste von ausgestorbenen Beutelthieren der Diluvialzeit (Diprotodon und Nototherium), welche weit größer als die größten noch lebenden Marsupialien waren. Diprotodon australis, dessen Schädel allein drei Fuß lang ist, übertraf das Flußpferd oder den Hippopotamus, dem es im Ganzen an schwerfälligem und plumpem Körperbau glich, noch an Größe.
Man kann diese ausgestorbene Gruppe, welche wahrscheinlich den riesigen placentalen Hufthieren der Gegenwart, den Flußpferden und Rhinoceros, entspricht, wohl als Hufbeutler (Barypoda) bezeichnen. Diesen sehr nahe steht die Ordnung der Känguruhs oder Springbeutler (Macropoda), die Sie alle aus den zoologischen Gärten kennen. Sie entsprechen durch die sehr verkürzten Vorderbeine, die sehr verlängerten Hinterbeine und den sehr starken Schwanz, der als Springstange dient, den Springmäusen unter den Nagethieren. Durch ihr Gebiß erinnern Sie dagegen an die Pferde, und durch ihre zusammengesetzte Magenbildung an die Wiederkäuer. Eine dritte Ordnung von pflanzenfressenden Beutelthieren entspricht durch ihr Gebiß den Nagethieren, und durch ihre unterirdische Lebensweise noch besonders den Wühlmäusen. Wir können dieselben daher als Nagebeutler oder wurzelfressende Beutelthiere (Rhizophaga) bezeichnen. Sie ist gegenwärtig nur noch durch das australische Wombat (Phascolomys) vertreten. Eine vierte und letzte Ordnung von pflanzenfressenden Beutelthieren endlich bilden die Kletterbeutler oder früchtefressenden Beutelthiere (Carpophaga), welche in ihrer Lebensweise und Gestalt theils den Eichhörnchen, theils den Affen entsprechen (Phalangista, Phascolarctus).
Die zweite Legion der Marsupialien, die fleischfressenden Beutelthiere (Zoophaga), zerfallen ebenfalls in vier Hauptgruppen oder Ordnungen. Die älteste von diesen ist die der Urbeutler oder insectenfressenden Beutelthiere (Cantharophaga). Zu dieser gehören wahrscheinlich die Stammformen der ganzen Legion, und vielleicht auch der ganzen Unterklasse. Wenigstens gehören alle stonesfielder Unterkiefer (mit Ausnahme des erwähnten Stereiognathus) insectenfressenden Beutelthieren an, welche in dem jetzt noch lebenden Myrmecobius ihren nächsten Verwandten besitzen. Doch war bei einem Theile jener oolithischen Urbeutler die Zahl der Zähne größer, als bei allen übrigen bekannten Säugethieren, indem jede Unterkieferhälfte von Thylacotherium 16 Zähne enthält (3 Schneidezähne, 1 Eckzahn, 6 falsche und 6 wahre Backzähne). Wenn in dem unbekannten Oberkiefer ebenso viele Zähne saßen, so hatte Thylacotherium nicht weniger als 64 Zähne, gerade doppelt so viel als der Mensch. Die Urbeutler entsprechen im Ganzen den Insectenfressern unter den Placentalthieren, zu denen Igel, Maulwurf und Spitzmaus gehören. Eine zweite Ordnung, die sich wahrscheinlich aus einem Zweige der ersteren entwickelt hat, sind die Rüsselbeutler oder zahnarmen Beutelthiere (Edentula), welche durch die rüsselförmig verlängerte Schnauze, das verkümmerte Gebiß und die demselben entsprechende Lebensweise an die Zahnarmen oder Edentaten {Tabelle Seite 468} {Tabelle Seite 469} unter den Placentalien, insbesondere an die Ameisenfresser erinnern.


Andrerseits entsprechen die Raubbeutler oder Raubbeutelthiere (Creophaga) durch Lebensweise und Bildung des Gebisses den eigentlichen Raubthieren oder Carnivoren unter den Placentalthieren. Es gehören dahin der Beutelmarder (Dasyurus) und der Beutelwolf (Thylacinus) von Neuholland. Obwohl letztere die Größe des Wolfes erreicht, ist er doch ein Zwerg gegen die ausgestorbenen Beutellöwen Ausstraliens (Thylacoleo), welche mindestens von der Größe des Löwen waren und Reißzähne von mehr als zwei Zoll Länge besaßen. Die achte und letzte Ordnung endlich bilden die Handbeutler oder die affenfüßigen Beutelthiere (Pedimana), welche sowohl in Australien als in Amerika leben. Sie finden sich häufig in zoologischen Gärten, namentlich verschiedene Arten der Gattung Didelphys, unter dem Namen der Beutelratten, Buschratten oder Opossum bekannt. An ihren Hinterfüßen kann der Daumen unmittelbar den vier übrigen Zehen entgegengesetzt werden, wie bei einer Hand, und sie schließen sich dadurch unmittelbar an die Halbaffen oder Prosimien untern den Placentalthieren an. Es wäre möglich, daß diese letzteren wirklich den Handbeutlern nächstverwandt sind und aus längst ausgestorbenen Vorfahren derselben sich entwickelt haben.
Die Genealogie der Beutelthiere ist sehr schwierig zu errathen, vorzüglich deßhalb, weil wir die ganze Unterklasse nur höchst unvollständig kennen, und die jetzt lebenden Marsupialien offenbar nur die letzten Reste des früheren Formenreichthums darstellen. Vielleicht haben sich die Handbeutler, Raubbeutler, und Rüsselbeutler als drei divergente Aeste aus der gemeinsamen Stammgruppe der Urbeutler entwickelt. In ähnlicher Weise sind vielleicht andrerseits die Nagebeutler, Springbeutler und Hufbeutler als drei auseinandergehende Zweige aus der gemeinsamen pflanzenfressenden Stammgruppe, den Kletterbeutlern hervorgegangen. Kletterbeutler aber und Urbeutler sind zwei divergente Aeste der gemeinsamen Stammform aller Beutelthiere, welche während der älteren Secundärzeit aus den Kloakenthieren entstand.
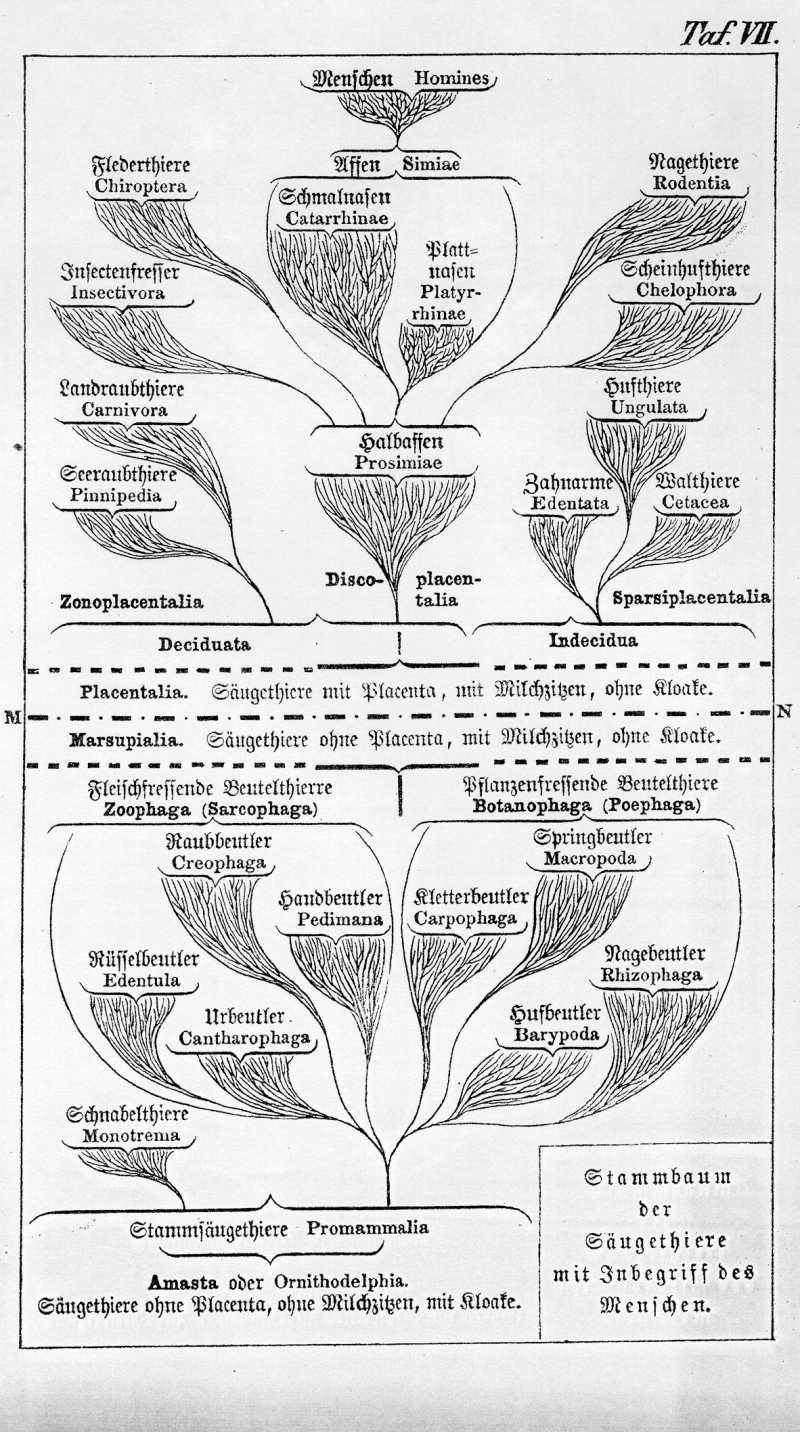
Die dritte und letzte Unterklasse der Säugethiere bilden die Placentalthiere oder Placentner (Monodelphia oder Placentalia). Sie ist bei weitem die wichtigste, umfangreichste und vollkommenste von den drei Unterklassen. Denn zu ihr gehören alle bekannten Säugethiere nach Ausschluß der Beutelthiere und Schnabelthiere. Auch der Mensch gehört dieser Unterklasse an und hat sich aus niederen Stufen derselben entwickelt.
Die Placentalthiere unterscheiden sich, wie ihr Name sagt, von den übrigen Säugethieren vor Allem durch den Besitz eines sogenannten Mutterkuchens oder Aderkuchens (Placenta). Das ist ein sehr eigenthümliches und merkwürdiges Organ, welches bei der Ernährung des im Mutterleibe sich entwickelnden Jungen eine höchst wichtige Rolle spielt. Die Placenta oder der Mutterkuchen (auch Nachgeburt genannt) ist ein weicher, schwammiger, rother Körper von sehr verschiedener Form und Größe, welcher zum größten Theile aus einem unentwirrbaren Geflecht von Adern oder Bluthgefäßen besteht. Ihre Bedeutung besteht in dem Stoffaustausch des ernährenden Blutes zwischen dem mütterlichen Fruchtbehälter oder Uterus und dem Leibe des Keimes oder Embryo (s. oben s. 243). Weder bei den Beutelthieren noch bei den Schnabelthieren ist dieses höchst wichtige Organ entwickelt. Von diesen beiden Unterklassen unterscheiden sich aber auch außerdem die Placentalthiere noch durch manche andere Eigenthümlichkeiten, so namentlich durch den Mangel der Beutelknochen, durch die höheren Ausbildung der inneren Geschlechtsorgane und durch die vollkommenere Entwickelung des Gehirns, namentlich des sogenannten Schwielenkörpers oder Balkens (Corpus callosum), welcher als mittlere Commissur oder Querbrücke die beiden Halbkugeln des großen Gehirns mit eineinander verbindet. Auch fehlt den Placentalien der eigenthümliche Hakenfortsatz des Unterkiefers, welcher die Beutelthiere auszeichnet. wie in diesen anatomischen Beziehungen der Beutelthiere zwischen den Gabelthieren und Placentalthieren in der Mitte stehen, wird Ihnen am besten durch nachfolgende Zusammenstellung der wichtigsten Charaktere der drei Unterklassen klar werden. {Tabelle S. 472 oben}

Die Placentalthiere sind in weit höherem Maaße mannichfaltig differenzirt und vervollkommnet, als die Beutelthiere, und man hat daher dieselben längst in eine Anzahl von Ordnungen gebracht, die sich hauptsächlich durch die Bildung des Gebisses und der Füße unterscheiden. Noch wichtiger aber, als diese, ist die verschiedenartige Ausbildung der Placenta und die Art ihres Zusammenhanges mit dem mütterlichen Fruchtbehälter. Bei den niederen drei Hauptordnungen der Placentalthiere nämlich, bei den Hufthieren, Walthieren und Zahnarmen, entwickelt sich zwischen dem mütterlichen und kindlichen Theil der Plancenta nicht jene eigenthümliche schwammige Haut, welche man als hinfällige Haut oder Decidua bezeichnet. Diese findet sich ausschließlich bei den sieben höher stehenden Ordnungen der Placentalthiere, und wir können diese letzteren daher nach Huxley in der Hauptgruppe der Decudualthiere (Deciduata) vereinigen. Diesen stehen die drei erstgenannten Legionen als Decidualose (Indecidua) gegenüber.
Die Placenta unterscheidet sich bei den verschiedenen Ordnungen der Placentalthiere aber nicht allein duch die wichtigen inneren Structurverschiedenheiten, welche mit dem Mangel oder der Anwesenheit einer Decidua verbunden sind, sondern auch durch die äußere Form des Mutterkuchens selbst. Bei den Indeciduen besteht derselbe meistens aus zahlreichen einzenen, zerstreuten Gefäßknöpfen oder Zotten, und man kann daher die Gruppe auch als Zottelplacentner (Sparsiplacentalia) bezeichnen.
Bei den Deciduaten dagegen sind die einzelnen Gefäßzotten zu einem zusammenhängenden Kuchen vereinigt, und dieser erscheint in zweierlei verschiedener Gestalt. In den einen nämlich umgiebt er den Embryo in Form eines geschlossenen Gürtels oder Ringes, so daß nur die beiden Pole der länglichrunden Eiblase von Zotten frei bleiben. Das ist der Fall bei den Raubthieren (Carnaria) und den Scheinhufern (Chelophora), die wir deshalb als Gürtelplacentner (Zonoplacentalia) zusammenfassen. In den anderen Deciduathieren dagegen, zu welchen auch der Mensch gehört, bildet die Placenta eine einfache runde Scheibe, und wir nennen sie daher Scheibenplacentner (Discoplacentalia). Das sind die fünf Ordnungen der Halbaffen, Nagethiere, Insectenfresser, Flederthiere und Affen, von welchen letzteren auch der Mensch im zoologischen Systeme nicht zu trennen ist.
Das die Placentalthiere erst aus den Beutelthieren sich entwickelt haben, darf auf Grund ihrer vergleichenden Anatomie und Entwickelungsgeschichte als ganz sicher angesehen werden, und wahrscheinlich fand diese höchst wichtige Entwickelung, die erste Entstehung der Placenta, erst im Beginn der Tertiärzeit, während der Anteocen-Periode, statt. Dagegen gehört zu den schwierigsten Fragen der thierischen Genealogie die wichtige Untersuchung, ob alle Placentalthiere aus einem oder aus mehreren getrennten Zweigen der Beutlergruppe entstanden sind, mit anderen Worten, ob die Entstehung der Placenta einmal oder mehrmal statt hatte. Als ich vor zwei Jahren in meiner generellen Morphologie zum ersten Male den Stammbaum der Säugethiere zu begründen versuchte, zog ich auch hier, wie meistens, die monophyletische oder einwurzelige Descendenzhypothese der polyphyletischen oder vielwurzeligen vor.
Ich nahm an, daß alle Placentner von einer einzigen Beutelthierform abstammten, die zum ersten Male eine Placenta zu bilden begann. Dann wären die Sparsiplacentalien, Zonoplacentalien und Discoplacentalien vielleicht als drei divergente Aeste einer gemeinsamen placentalen Stammform aufzufassen, oder man könnte auch denken, daß die beiden letzteren, die Deciduaten, sich erst später aus den Indeciduen entwickelt hätten, die ihrerseits unmittelbar aus den Beutlern entstanden seien. Jedoch giebt es andrerseits auch gewichtige Gründe für die andere Alternative, daß nämlich mehrere von Anfang verschiedene Placentnergruppen aus mehreren verschiedenen Beutlergruppen entstanden seien, daß also die Placenta selbst sich mehrmals unabhängig von einander gebildet habe. Dies ist unter anderen die Ansicht des ausgezeichnetsten englischen Zoologen, Huxley's. In diesem Falle wären zunächst als zwei ganz getrennte Gruppen vielleicht die Indeciduen und Decudaten aufzufassen.
Von den Indeciduen wäre möglicherweise die Ordnung der Hufthiere, als die Stammgruppe, aus den pflanzenfressenden Hufbeutlern oder Barypoden entstanden. Unter den Deciduaten dagegen würde vielleicht die Ordnung der Halbaffen, als gemeinsame Stammgruppe der übrigen Ordnungen, aus den Handbeutlern oder Pedimanen entstanden sein. Es wäre aber auch wohl möglich, daß die Deciduaten selbst wieder aus mehreren verschiedenen Beutler-Ordnungen entstanden seien, die Raubthiere z. B. aus den Raubbeutlern, die Nagethiere als den Nagebeutlern, die Halbaffen aus den Handbeutlern u. s. w. Da wir zur Zeit noch kein genügendes Erfahrungsmaterial besitzen, um diese äußerst schwierige Frage zu lösen, so lassen wir dieselbe auf sich beruhen, und wenden uns zur Geschichte der verschiedenen Placentner-Ordnungen, deren Stammbaum sich im Einzelnen oft in großer Vollständigkeit feststellen läßt.
Als die Stammgruppe der Decidualosen oder Zottenplacentner müssen wir, wie schon bemerkt, die Ordnung der Hufthiere (Ungulata) auffassen, aus welcher sich die beiden anderen Ordnungen, Walthiere und Zahnarme, wahrscheinlich erst später als zwei divergente Gruppen durch Anpassung an sehr verschiedene Lebensweise entwickelt haben. Doch sind die Zahnarmen oder Edentaten vielleicht auch ganz anderen Ursprungs.
Die Hufthiere gehören in vielen Beziehung zu den wichtigsten und interessantesten Säugethieren. Sie zeigen deutlich, wie uns das wahre Verständniß der natürlichen Verwandtschaft der Thiere niemals allein aus dem Studium der noch lebenden Formen, sondern stets nur durch gleichmäßige Berücksichtigung ihrer ausgestorbenen und versteinerten Blutsverwandten und Vorfahren erschlossen werden kann.
Wenn man in herkömmlicher Weise allein die lebenden Hufthiere berücksichtigt, so erscheint es ganz naturgemäß, dieselben in drei gänzlich verschiedene Ordnungen einzutheilen, nämlich
1. die Pferde oder Einhufer (Solidungula oder Equina);
2. die Wiederkäuer oder Zweihufer (Bisulca oder Ruminantia); und
3, die Dickhäuter oder Vielhufer (Multungula oder Pachyderma).
Sobald man aber die ausgestorbenen Hufthiere der Tertiärzeit mit in Betracht zieht, von denen wir sehr zahlreiche und wichtige Reste besitzen, so zeigt sich bald, daß jene Eintheilung, namentlich aber die Begrenzung der Dickhäuter, eine ganz künstliche ist, und daß diese drei Gruppen nur abgeschnittene Aeste des Hufthierstammes sind, welche durch ausgestorbenen Zwischenformen auf das engste verbunden sind. Die eine Hälfte der Dickhäuter, Nashorn, Tapir und Paläotherien zeigen sich auf das nächste mit den Pferden verwandt, und besitzen gleich diesen unpaarzehige Füße. Die andere Hälfte der Dickhäuter dagegen, Schweine, Flußpferde und Anoplotherien, sind durch ihre paarzehigen Füße viel enger mit den Wiederkäuern, als mit jenen ersteren verbunden. Wir müssen daher zunächst als zwei natürliche Hauptgruppen unter den Hufthieren die beiden Ordnungen der Paarhufer und der Unpaarhufer unterscheiden, welche sich als zwei divergente Aeste aus der alttertiären Stammgruppe der Stammhufer oder Prochelen entwickelt haben.
Die Ordnung der Unpaarhufer (Perissodactyla) umfaßt diejenigen Ungulaten, bei denen die mittlere (oder dritte) Zehe des Fußes viel stärker als die übrigen entwickelt ist, so daß sie die eigentliche Mitte des Hufes bildet. Es gehört hierher zunächst die uralte gemeinsame Stammgruppe aller Hufthieren, die Stammhufer (Prochela), welche schon in den ältesten eocenen Schichten versteinert vorkommt (Lophiodon, {Tabelle Seite 476} {Abbildung Seite 477} Pliolophus). An diese schließt sich unmittelbar derjenige Zweig derselben an, welcher die eigentliche Stammform der Unpaarhufer ist, die Paläotherien, welche fossil im oberen Eocen und unteren Miocen vorkommen. Aus den Paläotherien haben sich später als zwei divergente Zweige einerseits die Nashörner (Nasicornia) und Nashornpferde (Elasmotherida), andrerseits die Tapire, Lamatapire und Urpferde entwickelt. Die längst ausgestorbenen Urpferde oder Anchitherien vermittelten den Uebergang von den Paläotherien und Tapiren zu den Mittelpferden oder Hipparionen, die den noch lebenden echten Pferden schon ganz nahe stehen.

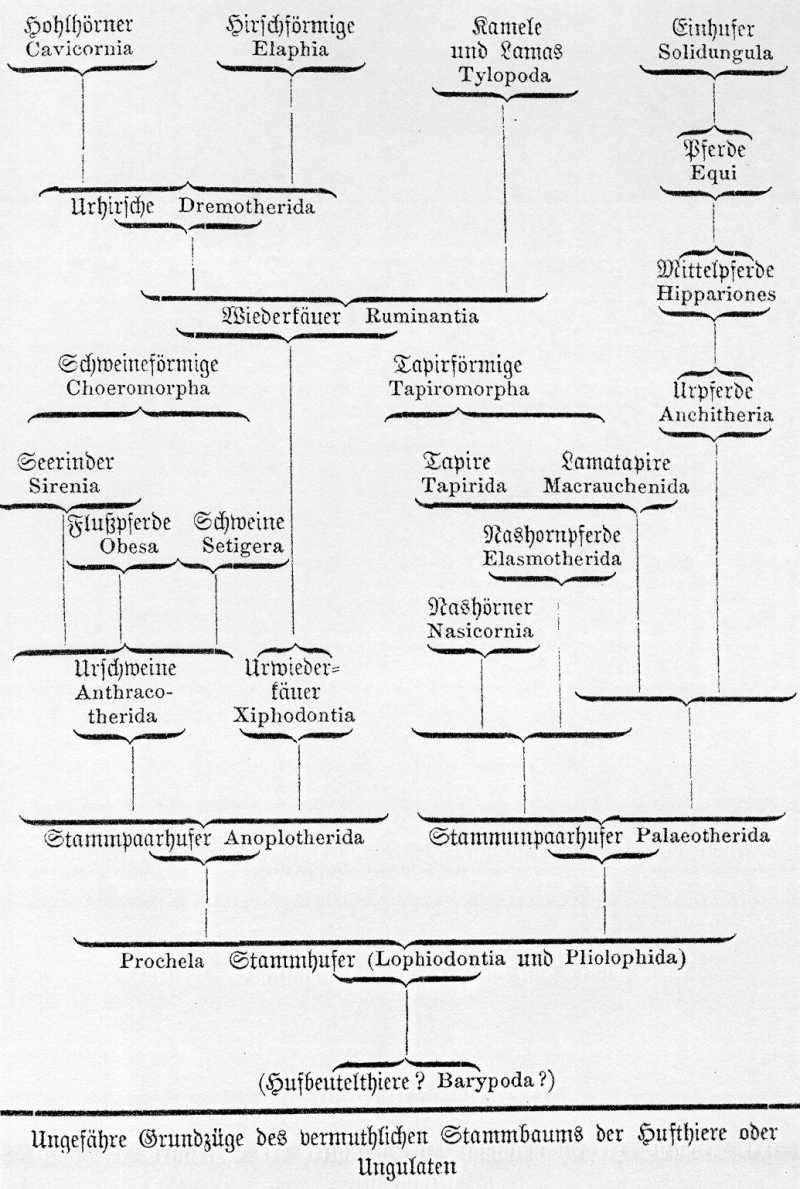
Die zweite Hauptgruppe der Hufthiere, die Ordnung der Paarhufer (Artiodactyla) enthält diejenigen Hufthiere, bei denen die mittlere (dritte) und die vierte Zehe des Fußes nahzu gleich stark entwickelt sind, so daß die Theilungsebene zwischen Beiden die Mitte des ganzen Fußes bildet. Sie zerfällt in die beiden Unterordnungen der Schweineförmigen und der Wiederkäuer. Zu den Schweineförmigen (Choeromorpha) gehört zunächst der andere Zweig der Stammhufer, die Anoplotherien, welche wir als die gemeinsame Stammform aller Paarhufer oder Artiodactylen betrachten. Aus dieser Stammgruppe entsprangen wahrscheinlich als zwei divergente Zweige einerseits die Urschweine oder Anthrakotherien, welche zu den Schweinen und Flußpferden, andrerseits die Xiphodonten, welche zu den Wiederkäuern hinüberführten.
Die ältesten Widerkäuer (Ruminantia) sind die Urhirsche oder Dremotherien, aus denen vielleicht als drei divergente Zweige die Hirschförmigen (Elaphia), die Hohlhörnigen (Cavicornia) und die Kamele (Tylopoda) sich entwickelt haben. Doch sind die letzteren in mancher Beziehung mehr den Unpaarhufern als den echten Paarhufern verwandt. Wie sich die zahlreichen Familien der Hufthiere dieser genealogischen Hypothese entsprechend gruppiren zeigt Ihnen die vorstehende systematische Uebersicht (S. 476). Aus Hufthieren, welche sich an das ausschließliche Leben im Wasser gewöhnten, und dadurch fischähnlich umbildeten, ist wahrscheinlich die merkwürdige Legion der Walthiere (Cetacea) entsprungen. Obwohl diese Thiere äußerlich manchen echten Fischen sehr ähnlich erscheinen, sind sie dennoch, wie schon Aristoteles erkannte, echte Säugethiere.
Durch ihren gesamten inneren Bau, sofern derselbe nicht durch Anpassung an das Wasserleben verändert ist, stehen sie den Hufthieren von allen übrigen bekannten Säugethieren am nächsten, und theilen namentlich mit ihnen den Mangel der Decidua und die zottenförmige Placenta. Noch heute bildet das Pflußpferd (Hippopotamus) eine Art von Uebergangsform zu den Seerindern (Serenia), und es ist demnach das wahrscheinlichste, daß die ausgestorbenen Stammformen der Cetaceen den heutigen Seerindern am nächsten standen, und sich aus Paarhufern entwickelten, welche dem Flußpferd verwandt waren.
Aus der Ordnung der pflanzenfressenden Walthiere (Phycoceta), zu welcher die Seerinder gehören, und welche demnach wahrscheinlich die Stammformen der Legion enthält, hat sich späterhin die andere Ordnung der fleischfressenden Walthiere (Sarcoceta) entwickelt. Von diesen letzteren sind die ausgestorbenen riesigen Zeuglodonten (Zeugloceta), deren fossile Skelete vor einiger Zeit als angebliche "Seeschlangen" (Hydrarchus) großes Aufsehen erregten, vermuthlich nur ein eigenthümlich entwickelter Seitenzweig der eigentlichen Walfische (Autoceta), zu denen außer den colossalen Bartenwalen auch die Potwale, Delphine, Narwale, Seeschweine u. s. w. gehören.
Die dritte und letzte Legion der Indeciduen oder Sparsiplacentalien bildet die seltsame Gruppe der Zahnarmen (Edentata). Sie ist aus den beiden Ordnungen der Scharrthiere und der Faulthiere zusammengesetzt. Die Ordnung der Scharrthiere (Effodientia) besteht aus den beiden Unterordnungen der Ameisenfresser (Vermilinguia, zu denen auch die Schuppenthiere gehören), und der Gürtelthiere (Cingulata), die früher durch die riesigen Glyptodonten vertreten waren. Die Ordnung der Faulthiere (Tardigrada) besteht aus den beiden Unterordnungen der kleinen jetzt noch lebenden Zwergfaulthiere (Bradypoda) und der ausgestorbenen schwerfälligen Riesenfaulthiere (Gravigrada).
Die ungeheuren versteinerten Reste dieser colossalen Planzenfresser deuten darauf hin, daß die ganze Legion im Aussterben begriffen und die heutigen Zahnarmen nur ein dürftiger Rest von den gewaltigen Edentaten der Diluvialzeit sind. Die nahen Beziehungen der noch heute lebenden Edentaten Südamerikas zu den ausgestorbenen Riesenformen, die sich neben jenen in demselben Erdtheil finden, machte auf Darwin bei seinem ersten Besuche Südamerikas einen solchen Eindruck, daß sie schon damals den Grundgedanken der Descendenztheorie in ihm anregten (S. oben S. 107). Uebrigens ist die Genealogie grade dieser Legion sehr schwierig. Vielleicht sind die Edentaten nichts weiter, als ein eigenthümlich entwickelter Seitenzweig der Ungulaten; vielleicht liegt aber auch ihre Wurzel ganz wo anders.
Wir verlassen nun die erste Hauptgruppe der Placentner, die Decidualosen, und wenden uns zur zweiten Hauptgruppe, den Deciduathieren (Deciduata), welche sich von jenen so wesentlich durch den Besitz einer hinfälligen Haut oder Decidua während des Embryolebens unterscheiden. Hier begegnen wir zuerst einer sehr einheitlich organisirten und natürlichen Gruppe, nämlich der Ordnung der Raubthiere (Carnaria). Sie werden wohl auch Gürtelplacentner (Zonoplacentalia) im engeren Sinne genannt, obwohl eigentlich gleicherweise die Scheinhufer oder Chelophoren diese Bezeichnung verdienen. Da aber diese letzteren im Uebrigen näher den Nagethieren als den Raubthieren verwandt sind, werden wir sie dort besprechen. Die Raubthiere zerfallen in zwei, äußerlich sehr verschiedene, aber innerlich nächst verwandte Unterordnungen, die Landraubthiere und die Seeraubthiere.
Zu den Landraubthieren (Carnivora) gehören die Bären, Hunde, Katzen u. s. w., deren Stammbaum sich mit Hilfe vieler ausgestorbener Zwischenformen annähernd errathen läßt. Zu den Seeraubthieren oder Robben (Pinnipedia) gehören die Seebären, Seehunde, Seelöwen, und als eigenthümliche angepaßte Seitenlinie die Walrosse oder Walrobben. Obwohl die Seeraubthiere äußerlich den Landraubthieren sehr unähnlich sehen, sind sie denselben dennoch durch ihren inneren Bau, ihr Gebiß und ihre eigenthümliche gürtelformige Placenta nächst verwandt und offenbar aus einem Zweige derselben, vermuthlich den Marderartigen (Mustelina) hervorgegangen.
Noch heute bilden unter den letzteren die Fischottern (Lutra) und noch mehr die Seeottern (Enhydris) eine unmittelbare Uebergangsform zu den Robben, und zeigen uns deutlich, wie der Körper der Landraubthiere durch Anpassung an das Leben im Wasser robbenähnlich umgebildet wird, und wie aus den Gangbeinen der ersteren die Ruderflossen der Seeraubthiere entstanden sind. Die letzteren verhalten sich demnach zu den ersteren ganz ähnlich, wie unter den Indeciduen die Walthiere zu den Hufthieren. In gleicher Weise wie das Flußpferd noch heute zwischen den extremen Zweigen der Rinder und der Seerinder in der Mitte steht, bildet dieSeeotter noch heute eine übriggebliebene Zwischenstufe zwischen den weit entfernten Zweigen der Löwen und der Seelöwen. Hier wie dort hat die gänzliche Umgestaltung der äußeren Körperform, welche durch Anpassung an ganz verschiedene Lebensbedingungen bewirkt wurde, die tiefe Grundlage der erblichen inneren Eigenthümlichkeiten nicht zu vermischen vermocht.
Von den übrigen Deciduaten (nach Ausschluß der Raubthiere) betrachte ich als gemeinsame Stammgruppe die Halbaffen (Prosimiae). Die merkwürdigen Thiere wurden bisher in einer und derselben Ordnung, die Blumenbach als Vierhänder (Quadrumana) bezeichnete, mit den Affen vereinigt. Indessen trenne ich sie von diesen gänzlich, nicht allein deshalb, weil sie von allen Affen viel mehr abweichen, als die verschiedenen Affen von einander, sondern auch, weil sie die interessantesten Uebergangsformen zu den übrigen Ordnungen der Deciduaten enthalten. Ich schließe daraus, daß die wenigen jetzt noch lebenden Halbaffen, welche überdies unter sich sehr verschieden sind, die letzten überlebenden Reste einer fast ausgestorbenen, einstmals formenreichen Stammgruppe darstellen, aus welcher sich alle übrigen Deciduaten (vielleicht mit der einzigen Ausnahme der Raubthiere und der Scheinhufer) als divergente Zweige entwickelt haben. Die alte Stammgruppe der Halbaffen selbst hat sich vermuthlich aus den Handbeutlern oder affenfüßigen Beutelthieren (Pedimana) entwickelt, welche in der Umbildung ihrer Hinterfüße zu einer Greifhand ihnen auffallend gleichen.
Die uralten (wahrscheinlich in der Anteocen-Periode entstandenen) Stammformen selbst sind natürlich längst ausgestorben, ebenso die allermeisten Uebergangsformen zwischen denselben und den übrigen Deciduaten-Ordnungen. Aber einzelne Reste der letzteren haben sich in den heute noch lebenden Halbaffen erhalten. Unter diesen bildet das merkwürdige Fingerthier von Madagaskar (Chiromys madagascariensis) den Rest der Leptodactylen-Gruppe und den Uebergang zu den Nagethieren. Der seltsame Pelzflatterer der Südsee-Inseln und Sunda-Inseln (Galeopithecus), das einzige Ueberbleibsel der Ptenopleuren- Gruppe, ist eine vollkommene Zwischenstufe zwischen den Halbaffen und den Flederthieren.
Die Langfüßer (Tarsius, Otolicnus) bilden den letzten Rest desjenigen Stammzweiges (Macrotarsi), aus dem sich die Insectenfresser entwickelten. Die Kurzfüßer endlich (Brachytarsi) vermitteln den Anschluß an die echten Affen. Zu den Kurzfüßern gehören die langschwänzigen Maki (Lemur), und die kurzschwänzigen Indri (Lichanotus) und Lori (Stenops), von denen namentlich die letzteren sich den vermuthlichen Vorfahren des Menschen unter den Halbaffen sehr nahe anzuschließen scheinen. Sowohl die Kurzfüßer als die Langfüßer leben weit zerstreut auf den Inseln des südlichen Asiens und Afrikas, namentlich auf Madagaskar, einige auch auf dem afrikanischen Festlande. Kein Halbaffe ist bisher lebend oder fossil in Amerika gefunden. Alle führen eine einsame, nächtliche Lebensweise und klettern auf Bäumen umher.
Unter den übrigen Deciduen-Ordnungen, welche wahrscheinlich alle von längst ausgestorbenen Halbaffen abstammen, ist auf der niedrigsten Stufe die formenreiche Ordnung der Nagethiere (Rodentia) stehen geblieben. Unter diesen stehen die Eichhornartigen (Sciuromorpha) den Fingerthieren am nächsten. Aus dieser Stammgruppe haben sich wahrscheinlich als zwei divergente Zweige die Mäuseartigen (Myomorpha) und die Stachelschweinartigen (Hystrichomorpha) entwickelt, von denen jene durch eocene Myoxiden diese durch eocene Psammoryctiden unmittelbar mit den Eichhornartigen zusammenhängen. Die vierte Unterordnung, die Hasenartigen (Lagomorpha), haben sich wohl erst später aus einer von jenen drei Unterordnungen entwickelt.
An die Nagethiere schließt sich sehr eng die merkwürdige Ordnung der Scheinhufer (Chelophora) an. Von diesen leben heutzutage nur noch zwei, in Asien und Afrika einheimische Gattungen, nämlich die Elephanten (Elephas) und die Klippdasse (Hyrax). Beide wurde bisher gewöhnlich zu den echten Hufthieren oder Ungulaten gestellt, mit denen sie in der Hufbildung der Füße übereinstimmen. Allein eine gleiche Umbildung der ursprünglichen Nägel oder Krallen zu Hufen findet sich auch bei echten Nagethieren, und gerade unter diesen Hufnagethieren (Subungulata), welche ausschließlich Südamerika bewohnen, finden sich neben kleineren Thieren (z. B. Meerschweinchen und Goldhasen) auch die größten aller Nagethiere, die gegen vier Fuß langen Wasserschweine (Hydrochoerus capybara).
Die Klippdasse, welche auch äußerlich den Nagethieren, namentlich den Hufnagern sehr ähnlich sind, wurden bereits früher von einigen berühmten Zoologen als eine besondere Unterordnung (Lamnungia) wirlich zu den Nagethieren gestellt. Dagegen betrachtete man die Elephanten, falls man sie nicht zu den Hufthieren rechnete, gewöhnlich als Vertreter einer besonderen Ordnung, welche man Rüsselthiere (Proboscidea) nannte. Nun stimmen aber die Elephanten und Klippdasse merkwürdig in der Bildung ihrer Placenta überein, und entfernen sich dadurch jedenfalls gänzlich von den Hufthieren. Allen es ist wohl denkbar, daß sich die gürtelförmige Placenta erst secundär aus der scheibenförmigen entwickelt hat. In diesem Falle könnte man daran denken, daß die Scheinhufer aus einem Zweige der Nagethiere, und ähnlich vielleicht die Raubthiere aus einem Zweige der Insectenfresser sich entwickelt haben. Jedenfalls stehen die Elephanten und die Klippdasse auch in anderen Beziehungen, namentlich in der Bildung wichtiger Skelettheile, der Gliedmaßen u. s. w., den Nagethieren und namentlich den Hufnagern, näher als den ächten Hufthieren.
Dazu kommt noch, daß mehrere ausgestorbene Formen, namentlich die merkwürdigen südamerikanischen Pfeilzähner (Toxodontia) in mancher Beziehung zwischen Elephanten und Nagethieren in der Mitte stehen. Daß die noch jetzt lebenden Elephanten und Klippdasse nur die letzten Ausläufer von einer einstmals formenreichen Gruppe von Scheinhufern sind, wird nicht allein durch die sehr zahlreichen versteinerten Arten von Elephant und Mastodon bewiesen (unter denen manche größer, manche aber auch viel kleiner, als die jetzt lebenden Elephanten sind), sondern auch durch die merkwürdigen miocenen Dinotherien (Gonyognatha), zwischen denen und den nächstverwandten Elephanten noch eine lange Reihe von unbekannten verbindenden Zwischenformen liegen muß. Alles zusammengenommen ist heutzutage die wahrscheinlichste von allen Hypothesen, die man sich über die Entstehung und die Verwandtschaft der Elephanten, Dinotherien, Toxodonten und Klippdassen bilden kann, daß dieselben die letzten Ueberbleibsel einer formenreichen Gruppe von Scheinhufern sind, die sich aus den Nagethieren, und zwar wahrscheinlich aus Subungulaten, entwickelt hatte.
Die Ordnung der Insectenfresser (Insectivora) hat sich wahrscheinlich aus Halbaffen entwickelt, welche den heute noch lebenden Langfüßern (Macrotarsi) nahe standen. Sie spaltet sich in zwei Ordnungen, Menotyphla und Lipotyphla. Von diesen sind die älteren wahrscheinlich die Menotyphlen, welche sich durch den Besitz eines Blinddarms oder Typhlon von den Lipotyphlen unterscheiden. Zu den Menotyphlen gehören die kletternden Tupajas der Sunda-Inseln und die springenden Makroscelides Afrikas. Die Lipotyphlen sind bei uns durch die Spitzmäuse, Maulwürfe und Igel vertreten. Durch Gebiß und Lebensweise schließen sich die Insectenfresser mehr den Raubthieren, durch die scheibenförmige Placenta und die großen Samenblasen dagegen mehr den Nagethieren an.
Den Insectenfressern sehr nahe steht die merkwürdige Ordnung der fliegenden Säugethiere oder Flederthiere (Chiroptera). Sie hat sich durch Anpassung an fliegende Lebensweise in ähnlicher Weise auffallend umgebildet, wie die Seeraubthiere und die Walthiere durch Anpassung an schwimmende Lebensweise. Wahrscheinlich hat auch diese Ordnung ihre Wurzel in den Halbaffen, mit denen sie noch heute durch die Pelzflatterer (Galeopithecus) eng verbunden ist. Von den beiden Unterordnungen der Flederthiere haben sich wahrscheinlich die insectenfressenden oder Fledermäuse (Nycterides) erst später aus den früchtefressenden oder Flederhunden (Pterocynes) entwickelt; denn die letzteren stehen in mancher Beziehung den Halbaffen noch näher als die ersteren. Als letzte Säugethierordnung hätten wir nun endlich noch die echten Affen (Simiae) zu besprechen. Da aber im zoologischen System zu dieser Ordnung auch das Menschengeschlecht gehört, und da dasselbe sich aus einem Zweige dieser Ordnung ohne allen Zweifel historisch entwickelt hat, so wollen wir die genauere Untersuchung ihres Stammbaumes und ihrer Geschichte einem besonderen Vortrage vorbehalten.
Stammbaum und Geschichte des Thierreichs

 Ursprung und Stammbaum des Menschen
Ursprung und Stammbaum des Menschen
Renaissance Forum - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - erstes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Jacob Burckhardt - Die Kultur der Renaissance in Italien. Ein Versuch
Leonardo da Vinci Wissenschaftler - Erfinder - Künstler
Venedig - Eine Liebeserklärung an eine Stadt
William Shakespeare animiert (wahrscheinlich oder zufällig...2/3 zu 1/3) William Turner
Philosophie für Schnelldenker - Besinnliche Philosophie
Startseite - © Copyright 2004- - Susanne Albers - Kiehlufer 125 - D 12059 Berlin