Startseite - Impressum - Datenschutzerklärung - News - Ich diene dem, der hört - https://offenunddirekt.de - Mein Gratis-Coaching für Dich - Kraft tanken - Spirituelles - Kunst - Technik - Literatur - Kultur - Politik - Architektur - Spaß - Psychologie - Spiele - Fotos + Animationen - Gästebuch - Persönliches - Blog - Die Stimme Gottes - mein Youtube Kanal - erstes Rätsel - Renaissance Forum - Chat - Quiz - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - Zufallsspiel - letztes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Ernst Haeckel - Die Natürliche Schöpfungsgeschichte 1868
Siebzehnter Vortrag.
Stammbaum und Geschichte des Thierreichs.
I. Stammbaum und Geschichte der wirbellosen Thiere.
(Hierzu Taf. III, IV und V.)
Meine Herren! Das natürliche System der Organismen, welches wir ebenso im Thierreich wie im Pflanzenreich zunächst als Leitfaden für unsere genealogischen Untersuchungen benutzen müssen, ist hier wie dort erst neueren Ursprungs, und wesentlich durch die Fortschritte unseres Jahrhunderts in der vergleichenden Anatomie und Ontogenie bedingt. Die Klassificationsversuche des vorigen Jahrhunderts bewegten sich fast sämmtlich noch in der Bahn des künstlichen Systems, welches zuerst Carl Linné in strengerer Form aufgestellt hatte. Das künstliche System unterscheidet sich von dem natürlichen wesentlich dadurch, daß es nicht die gesammte Organisation und die innere, auf der Blutsverwandtschaft beruhende Formverwandtschaft zur Grundlage der Eintheilung macht, sondern nur einzelne und dazu meist noch äußerliche, leicht in die Augen fallende Merkmale. So unterschied Linné seine 24 Klassen des Pflanzenreichs wesentlich nach der Zahl, Bildung und Verbindung der Staubgefäße. Ebenso unterschied derselbe im Thierreiche sechs Klassen wesentlich nach der Beschaffenheit des Herzens und des Blutes.
Diese sechs Klassen waren:
1. die Säugethiere;
2. die Vögel;
3. die Amphibien;
4. die Fische;
5. die Insecten und
6. die Würmer.
Diese sechs Thierklassen Linné's sind aber keineswegs von gleichem Werthe, und es war schon ein wichtiger Fortschritt, als Lamarck zu Ende vorigen Jahrhunderts die vier ersten Klassen als Wirbelthiere (Vertebrata) zusammenfaßte, und diesen die übrigen Thiere, die Insecten und Würmer Linné's, als eine zweite Hauptabtheilung, als Wirbellose (Invertebrata) gegenüberstellte. Eigentlich griff Lamarck damit auf den Vater der Naturgeschichte, auf Aristoteles zurück, welche diese beiden großen Hauptgruppen bereits unterschieden, und die ersten Blutthiere, die letzteren Blutlose genannt hatte.
Den nächsten großen Fortschritt zum natürlichen System des Thierreichs thaten einige Decennien später zwei der verdienstvollsten Zoologen, Carl Ernst Bär und George Cuvier. Wie schon früher erwähnt wurde, stellten dieselben fast gleichzeitig, und uabhängig von einander, die Behauptung auf, daß mehrere grundverschiedene Hauptgruppen im Thierreich zu unterscheiden seien, von denen jede einen ganz eigenthümlichen Bauplan oder Typus besitze. (Vergl. oben S. 42, 43).
In jeder dieser Hauptabtheilungen giebt es eine baumartig verzweigte Stufenleiter von sehr einfachen und unvollkommenen bis zu höchst zusammengesetzten und entwickelten Formen. Der Ausbildungsgrad innerhalb eines jeden Typus ist ganz unabhängig von dem eigenthümlichen Bauplan, der dem Typus als besonderer Charakter zu Grunde liegt. Dieser "Typus" wird durch das eigenthümliche Lagerungsverhältniß der wichtigsten Körpertheile und die Verbindungsweise der Organe bestimmt. Der Ausbildungsgrad dagegen ist abhängig von der mehr oder weniger weitgehenden Arbeitstheilung oder Differenzirung der Plastiden und Organe. Diese außerordentlich wichtige und fruchtbare Idee begründete Bär, welcher sich auf die individuelle Entwickelungsgeschichte der Thiere stützte, viel klarer und tiefer als Cuvier, welcher sich bloß an die Resultate der vergleichenden Anatomie hielt. Doch erkannte weder dieser noch jener die wahre Ursache jenes merkwürdigen Verhältnisses. Diese wird uns erst durch die Descendenztheorie enthüllt. Sie zeigt uns, daß der gemeinsame Typus oder Bauplan durch die Vererbung, der Grad der Ausbildung oder Sonderung dagegen durch die Anpassung bedingt ist. (Gen. Morph. II, 10).
Sowohl Bär als Cuvier unterscheiden im Thierreich vier verschiedene Typen oder Baupläne und theilten dasselbe dem entsprechend in vier große Hauptabtheilungen (Zweige oder Kreise) ein.
Die erste von diesen wird durch die Wirbelthiere (Vertebrata) gebildet, welche die vier ersten Klassen Linné's umfassen: die Säugethiere, Vögel, Amphibien und Fische.
Den zweiten Typus bilden die Gliederthiere (Articulata), welche die Insecten Linné's, also die eigentlichen Insecten, die Tausendfüße, Spinnen und Krebse, außerdem aber auch einen großen Theil der Würmer, insbesondere die gegliederten Würmer enthalten.
Die dritten Hauptabtheilung umfaßt die Weichthiere (Mollusca): die Pulpen, Schnecken, Muscheln, und einige verwandte Gruppen.
Der vierte und letzte Kreis des Thierreichs endlich ist aus den verschiedenen Strahlthieren (Radiata) zusammengesetzt, welche sich auf den ersten Blick von den drei vorhergehenden Typen durch ihre "strahlige", blumenähnliche Körperform unterscheiden.
Während nämlich bei den Weichthieren, Gliederthieren und Wirbelthieren der Körper aus zwei symmethrisch-gleichen Seitenhälften besteht, aus zwei Gegenstücken oder Antimeren, von denen das eine das Spiegelbild des anderen darstellt, so ist dagegen bei den sogenannten Strahlthieren der Körper aus mehr als zwei, gewöhnlich vier, fünf oder sechs Gegenstücken zusammengsetzt, welche wie bei einer Blume um eine gemeinsame Hauptaxe gruppirt sind. So auffallend dieser Unterschied zunächst auch erscheint, so ist er doch im Grunde nur von höchst untergeordneter Bedeutung.
Die Aufstellung dieser natürlichen Hauptgruppen, Typen oder Kreise des Thierreichs, durch Bär und Cuvier war der größte Fortschritt in der Klassification seit Linné. Die drei Gruppen der Wirbelthieren, Gliederthiere und Weichthiere sind so naturgemäß, daß sie noch heutzutage fast allgemein beibehalten werden.
Dagegen mußte die unnatürliche Vereinigung der Strahlthiere bei genauerer Erkenntniß alsbald aufgelöst werden, und dieser wichtige Fortschritt wurde 1848 durch Leuckart gethan.
Er wies zuerst nach, daß darunter zwei grundverschiedene Typen vermischt seien, nämlich einerseits die Sternthiere (Echinodermata): die Seesterne, Seelilien, Seeigel und Seewalzen; andrerseits die Pflanzenthiere (Coelenterata): die Schwämme, Korallen, Schirmquallen und Kammquallen. Gleichzeitig wurden durch Siebold die Infusionsthierchen oder Infusorien mit den Wurzelfüßern oder Rhizopoden in einer besonderen Hauptabtheilung des Thierreichs als Urthiere (Protozoa) vereinigt. Dadurch stieg die Zahl der thierischen Typen oder Kreise auf sechs. Endlich wurde dieselbe noch dadurch um einen siebenten Typus vermehrt, daß die meisten neueren Zoologen die Hauptabtheilung der Gliederthiere oder Articulaten in zwei Gruppen trennten, einerseits die mit gegliederten Beinen versehenen Gliedfüßer (Arthropoda) welche den Insecten im Sinne Linné's entsprechen, nämlich die eigentlichen (sechsbeinigen) Insecten, die Tausendfüße, Spinnen und Krebse; andrerseits die fußlosen oder mit ungegliederten Füßen versehenen Würmer (Vermes). Diese letzteren umfassen nur die eigentlichen oder echten Würmer (die Ringelwürmer, Rundwürmer, Plattwürmer u. s. w.), und entsprechen daher keineswegs den Würmern in Linné's Sinne,welcher dazu auch noch die Weichthiere, Strahlthiere und viele andere rechnete.
So wäre denn nach der Anschauung der neueren Zoologen, welche Sie fast in allen Hand- und Lehrbüchern der gegenwärtigen Thierkunde vertreten finden, das Thierreich aus sieben ganz verschiedenen Hauptabtheilungen oder Typen zusammengesetzt, deren jede durch einen charakteristischen, ihr ganz eigenthümlichen sogenannten Bauplan ausgezeichnet, und von jeder der anderen völlig verschieden ist. In dem natürlichen System des Thierreichs, welches ich Ihnen jetzt als den wahrscheinlichen Stammbaum desselben entwickeln werde, schließe ich mich im Großen und Ganzen dieser üblichen Eintheilung an, jedoch nicht ohne einige Modificationen, welche ich in Betreff der Genealogie für sehr wichtig halte. Unverändert in ihrem bisherigen Umfange werde ich die drei Typen der Wirbelthiere, Gliedfüßer, und Sternthiere beibehalten. Dagegen müssen die drei Gruppen der Weichthiere, Würmer und Pflanzenthiere einige Veränderungen ihres Gebiets erleiden. Den siebenten und letzten Kreis, den der Urthiere oder Protozoen, löse ich ganz auf. Den größten Theil der jetzt gewöhnlich als Urthiere angesehenen Organismen, nämlich die Wurzelfüßer, Amoeboiden, Geißelschwärmer und Meerleuchten betrachte ich als Protisten und habe Ihnen dieselben bereits vorgeführt. Von den beiden noch übrigen Klassen der Urthiere betrachte ich die Schwämme als Wurzel des Pflanzenthierstammes, die Infusorien als Wurzel des Würmerstammes.
Die sechs Zweige oder Kreise des Thierreichs, welche nach Ausscheidung der Protozoen übrig bleiben, sind ohne Zweifel durch ihre Anatomie und Entwickelungsgeschichte dergestalt charakterisirt, daß man sie im Sinne von Bär und Cuvier als selbstständige "Typen" auffassen kann. Trotz aller Mannichfaltigkeit in der äußeren Form, welche innerhalb jedes dieser Typen sich entwickelt, ist dennoch die Grundlage des inneren Baues, das wesentliche Lagerungsverhältniß der Körpertheile, welches den Typus bestimmt, so constant, bei allen Gliedern jedes Typus so übereinstimmend, daß man dieselben eben wegen dieser inneren Formverwandtschaft im natürlichen System in einer einzigen Hauptgruppe vereinigen muß. Daraus folgt aber unmittelbar, daß diese Vereinigung auch im Stammbaum des Thierreichs stattfinden muß. Denn die wahre Ursache jener innigen Formverwandtschaft kann nur die wirkliche Blutsverwandtschaft sein. Wir können also ohne Weiteres den wichtigen Satz aufstellen, daß alle Thiere welche zu einem und demselben Kreis oder Typus gehören, von einer und derselben ursprünglichen Stammform abstammen müssen. Mit anderen Worten, der Begriff des Kreises oder Typus, wie er in der Zoologie seit Bär und Cuvier für die wenigen obersten Hauptgruppen oder "Unterreiche" des Thierreichs gebräuchlich ist, fällt zusammen mit dem Begriffe des Stammes oder Phylum, wie ihn die Descendenztheorie für die Gesammheit derjenigen Organismen anwendet, welche ohne Zweifel blutsverwandt sind, und eine gemeinsame Wurzel besitzen.
Die übereinstimmenden Zeugnisse der vergleichenden Anatomie, Embryologie und Paläontologie begründen diese Blutsverwandtschaft aller Angehörigen eines jeden Typus so sicher, daß schon jetzt darüber kaum ein Zweifel herrschen kann. Wenigstens gilt dies fast ohne Widerspruch von den fünf Stämmen der Wirbelthiere, Gliedfüßer, Weichthiere, Sternthiere und Pflanzenthiere. Zweifelhafter ist dies bei den Würmern, deren Kreis auch in seiner heutigen Zusammensetzung immer noch ein buntes Gemisch von sehr verschiedenartigen Thieren darstellt, welche wesentlich nur in negativen Merkmalen, in der tiefen Stufe ihre Organisation und in dem indifferenten Charakter ihres Baues übereinstimmen. Noch heute ist ebenso wie zu Zeiten Linné's die Würmerklasse die allgemeine Rumpelkammer der Zoologie, in welche die Systematiker alle Thiere hineinwerfen, die sie in keinem anderen Typus oder Phylum mit Sicherheit unterbringen können. Dieses seltsame Verhältniß hat aber seinen guten Grund, und zwar darin, daß wir mit größter Wahrscheinlichkeit den Würmerstamm (in seinem heutigen Umfang) als die gemeinsame Wurzel oder Stammgruppe des ganzen Thierreichs ansehen können.
Obwohl jeder der fünf Stämme (nach Ausschluß des Würmerstammes) eine aufsteigende baumförmig verzweigte Stufenleiter von sehr einfachen und niederen zu sehr zusammengesetzten und hochorganisirten Thieren darstellt, so sind dennoch die unvollkommensten und niedersten Formen derselben immer bereits so differenzirt, daß sie nicht die ursprünglichen Stammformen des ganzen Stammes darstellen können. Dies gilt ebenso von den niedersten Stufen der Wirbelthiere und Gliedfüßer, wie von den unvollkommensten Formen der Weichthiere, Sternthiere und Pflanzenthiere. Wollen wir aber die ersten und ältesten Vorfahren derselben erkennen, so müssen wir nothwendig auf noch tiefer stehende Organismen zurückgehen.
Die Embryologie der Thiere belehrt uns, daß jedes Individuum sich aus einer einfachen Zelle, einem Ei entwickelt, und hieraus können wir, auf den innigen ursächlichen Zusammenhang zwischen Ontogenie und Phylogenie gestützt, unmittelbar den wichtigen Schluß ziehen, daß auch die ältesten Stammformen eines jeden Phylum einfache Zellen, gleich den Eiern, waren. Diese Zellen selbst aber müssen, wie ich Ihnen schon früher zeigte, von Moneren abstammen, die durch Urzeugung entstanden sind. Welche Formenkette liegt nun aber zwischen jenen einfachen Stammzellen und zwischen den verhältnißmäßig schon hoch organisirten Thieren, die wir heutzutage als die niedersten und ältesten Formen eines jeden der fünf genannten Stämme ansehen?
Auf diese Frage erhalten wir durch die vergleichende Anatomie und Embryologie zwar keine ganz bestimmte Antwort, aber doch einen sehr wichtigen Hinweis. Es zeigt sich nämlich, daß unter der bunten Formenmasse des gestaltenreichen Würmerstammes eine ganze Anzahl von interessanten Thierformen versteckt ist, welche wir mit einem mehr oder weniger hohen Grade von Wahrscheinlichkeit als Uebergangsformen von den niederen Würmern zu den niedersten Entwickelungsstufen der fünf übrigen Stämme ansehne können.
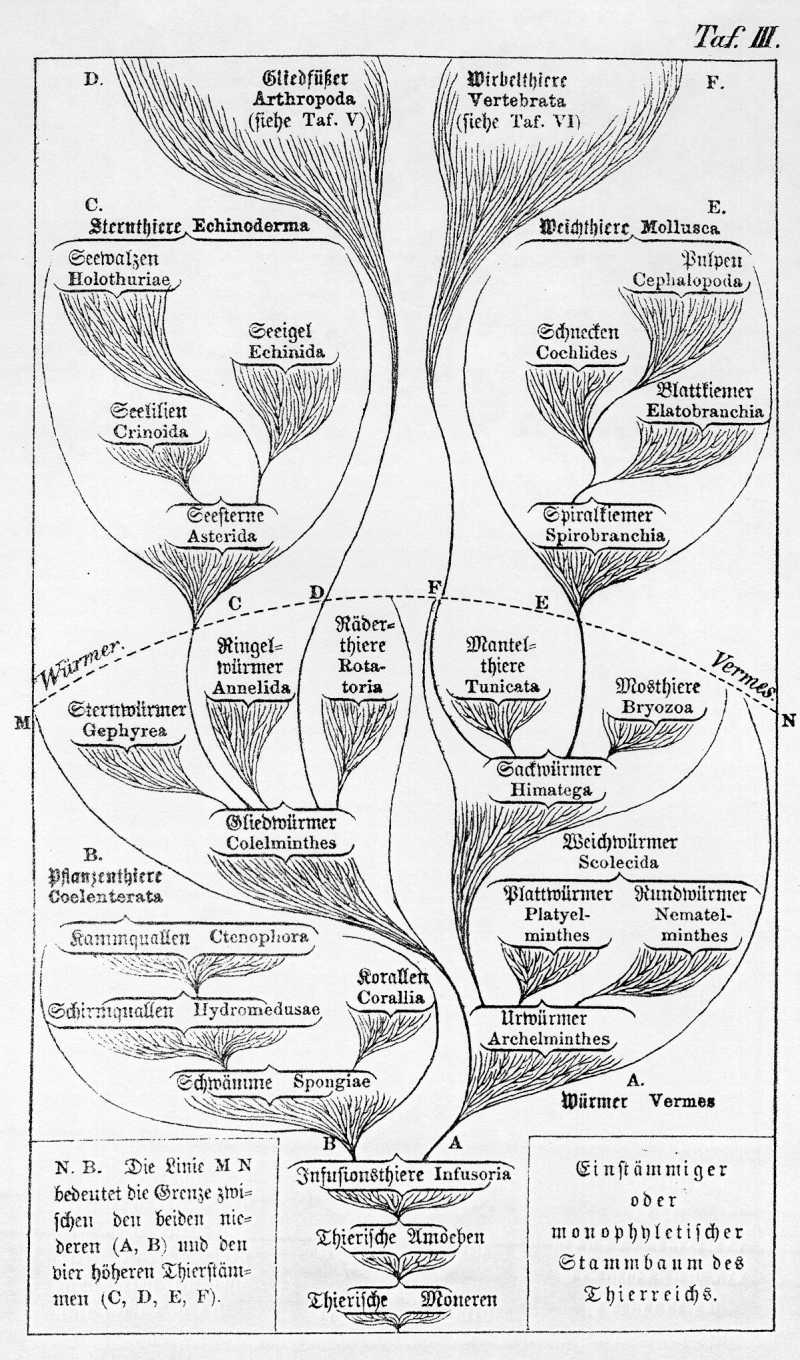
Wir dürfen in ihnen noch jetzt lebende nahe Verwandte von jenen längst ausgestorbenen Würmern vermuthen, aus denen sich in altersgrauer primordialer Vorzeit die fünf Stammformen der fünf übrigen Phylen entwickelten. So gleichen namentlich einige Infusionsthiere den ersten Jugendzuständen der Pflanzenthiere. Einige Weichwürmer und die Mosthiere schließen sich an die Weichthiere an. Die Sternwürmer und einige Ringelwürmer führen uns zu den Sternthieren hinüber, andere Ringelwürmer dagegen und die Räderthiere zu den Gliederfüßern. Die Mantelthiere endlich schließen sich zunächst an die Wirbelthiere an, indem die Jugendzustände von den niedersten Formen beider Gruppen nahe verwandt sind. Erwägen wir nun einerseits diese unleugbare anatomische und embryologische Verwandtschaft einzelner Würmergruppen mit den niedersten und tiefststehenden Ausgangsformen der fünf übrigen Stämme, andrerseits die vielfache verwandtschaftliche Verketten, durch welche die verschiedenen Gruppen des Würmerstammes trotz aller Verschiedenheiten unter sich innig verbunden sind, so gelangen wir schließlich zu der Anschauung, daß auch für das gesammte Thierreich ein gemeinsamer Ursprung aus einer einzigen Wurzel oder Stammform das Wahrscheinlichste ist.Auch hier, wie im Pflanzenreich, gewinnt bei näherer und eingehenderer Betrachtung die einstämmige oder monophyletische Descendenzhypothese, wie sie auf Taf. III. dargestellt ist, das Uebergewicht über die entgegengesetzte, vielstämmige oder polyphyletische Hypothese, von welcher Ihnen die nachstehende Tabelle (S. 392) eine Anschauung giebt.
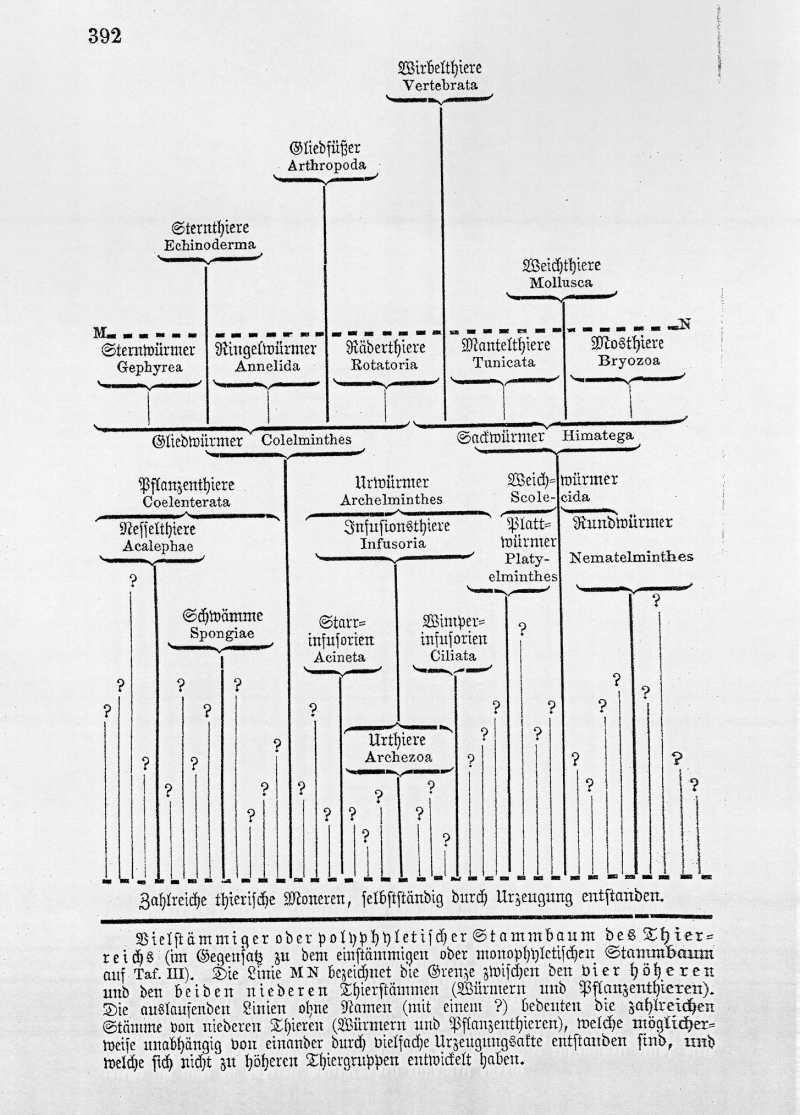
Die polyphyletische Hypothese vom Ursprung des Thierreichs kann in sehr verschiedener Form gedacht werden. Im Gegensatz zu der auf S. 392 dargestellten Form derselben könnte man es zunächst z. B. für das Wahrscheinlichste halten, daß jeder der sechs thierischen Stämme selbstständigen Ursprungs ist und sich ganz unabhängig von den fünf anderen aus einer besonderen Zellenform entwickelt hat, die von einem besonderen, durch Urzeugung entstandenen Moner abstammt. Gegen diese Vorstellung spricht erstens die merkwürdige Uebereinstimmung der frühesten embryonalen Entwickelungszustände bei den verschiedenen Stämmen, und zweitens die Menge von verbindenden Uebergangsformen, welche einerseits zwischen den verschiedenen Gruppen des Würmerstammes, und andrerseits zwischen diesen und den niedersten, auf tiefster Sonderungsstufe stehen gebliebenen Thieren der fünf übrigen Stamme existiren.
Die wahrscheinlichste genealogische Hypothese über den Ursprung und die paläontologische Entwickelung des Thierreichs ist demnach folgende (Taf. III). Durch Urzeugung entstanden zuerst thierische Moneren, gleich denen des Pflanzenreichs und des Protistenreichs ganz einfache structurlose Plasmastücke, aber von beiden durch leichte Unterschiede in der chemischen Zusammensetzung ihres eiweißartigen Plasma, und durch die daraus folgende Entwickelung zu echt thierischen Formen sich unterscheidend. Indem im Inneren dieser gleichartigen Moneren sich ein Kern von dem umgebenden Protoplasma sonderte, entstanden die ersten thierischen Zellen, ebenfalls nicht in ihrer Form, sondern nur in ihrer chemischen Zusammensetzung von den einfachsten selbstständigen Zellen unter den Urpflanzen und Protisten verschieden.
Diese nackten einzelligen Thiere, an Form gleichwerthig den Eiern der vielzelligen Thiere, lebten anfangs selbstständig, gleich den heute noch lebenden Amoeben. Später aber bildeten sie, in Colonien beisammen bleibend, vielzellige Körper, gleich dem kugeligen Haufen von Furchungskugeln, welcher bei den vielzelligen Thieren aus der wiederholten Theilung des Eies entsteht (Vergl. Fig. 2, S. 145, und Fig. 3, 4, S. 146). Aus diesen einfachen Haufen gleichartiger Zellen gingen allmählich durch Sonderung und Vervollkommnung die niedersten Würmer hervor, welche in den heute noch lebenden Infusionsthierchen ihre nächsten Verwandten besitzen. Die Ontogenie vieler Würmer, ferner vieler Pflanzenthiere, Sternthiere und Weichthiere, wiederholt uns noch heutzutage jenen wichtigen Vorgang der Phylogenie, indem das gefurchte Ei, d. h. der vielzellige, aus der Eitheilung entstandene Körper sich zunächst in einen bewimperten "infusorienartigen" Embryo oder Larve verwandelt.
Aus gleichen bewimperten Infusorien entstanden dann durch weitere Differenzirung die {Abbildung Seite 392} Vielstämmiger oder polyphyletischer Stammbaum des Thierreichs (im Gegensatz zu dem einstämmigen oder monophyletischen Stammbaum auf Taf. III). Die Linie M N bezeichnet die Grenze zwischen den vier höheren und den beiden niederen Thierstämmen (Würmern und Pflanzenthieren). Die auslaufenden Linien ohne Namen (mit einem ?) bedeuten die zahlreichen Stämme von niederen Thieren (Würmern und Pflanzenthieren), welche möglicherweise unabhängig von einander durch vielfache Urzeugungsakte entstanden sind, und welche sich nicht zu höheren Thiergruppen entwickelt haben. {Abbildung Seite 393 } niedersten Formen der bewimperten Strudelwürmer oder Turbellarien, Weichwürmer, welche wir als die gemeinsame Stammgruppe aller übrigen Würmerklassen ansehen können. Viele von den letzteren blieben bis auf den heutigen Tag auf der niedersten Entwickelungsstufe des Wurmes stehen. Einige wenige aber entwickelten sich nach verschiedenen Richtungen hin zu höheren Formen, welche die Stammformen für die übrigen, höheren Thierstämme wurden.
Wenn diese Hypothese, wie ich glaube, richtig ist, so würden die sechs Stämme des Thierreichs in genealogischer Beziehung keineswegs gleichwerthig sein. (Vergl. Taf. III.) Denn der Würmerstamm würde dann als die gemeinsame Stammgruppe der fünf übrigen Stämme zu betrachten sein. Diese letzteren verhielten sich unter einander wie fünf Geschwister, welche in dem ersteren ihre gemeinsame Elternform haben. Unter den fünf Geschwisterstämmen selbst würden wir aber wieder den Stamm der Pflanzenthiere oder Coelenteraten in sofern den vier übrigen entgegenstellen müssen, als der erstere einen viel geringeren Grad der Blutsverwandtschaft zu den echten Würmern offenbart, als die vier letzteren.
Wahrscheinlich hat sich der erstere in viel früherer Primordialzeit bereits von den tiefsten Stufen des Wurmstammes, von den Urwürmern oder Infusorien abgezweigt und selbstständig entwickelt, während die Stammformen der vier übrigen Stämme noch gar nicht von echten Würmern zu trennen waren. Diese letzteren haben sich wohl erst in viel späterer Zeit von den Würmern gesondert, als der Würmertypus längst die niedere und indifferente Stufe der Urwürmer überschritten hatte. Selbst wenn wir für die vier Stämme der Wirbelthiere, Gliedfüßer, Weichthiere und Sternthiere mit Bestimmtheit einen gemeinsamen Ursprung aus verschiedenen Zweigen des einheitlichen Würmerstammes annehmen, können wir doch über die Abstammung der Pflanzenthiere von den Würmern noch sehr in Zweifel bleiben, weil eben die niedersten Formen der letzteren, aus denen die ersten Pflanzenthiere entsprungen sein müßten, nur ganz indifferente und vielleicht ganz selbstständig entstandene Urwürmer gewesen sein können. Ich werde diesen genealogischen Bedenken in der nachfolgenden Entwickelung des thierischen Stammbaums dadurch einen berechtigten Ausdruck geben, daß ich die Pflanzenthiere als eine eigene, von den übrigen Thierstämmen entferntere Gruppe voranstelle, und auf diese erst die Würmer folgen lasse, aus denen sich die vier höheren Stämme des Thierreichs entwickelt haben.
Bevor ich nun diese Aufgabe in Angriff nehme und Ihnen meine genealogische Hypothese von der historischen Entwickelung der Thierstämme näher erläutere, wird es zweckmäßig sein, wie wir schon vorher beim Pflanzenreiche gethan haben, das ganze "natürliche System" des Thierreichs in einer Tabelle übersichtlich zusammen zu stellen, und die Hauptklassen und Klassen zu nennen, welche wir in jedem der sechs großen Thierstämme unterscheiden. Die Zahl dieser obersten Hauptabtheilungen ist im Thierreiche viel größer als im Pflanzenreiche, schon aus dem einfachen Grunde, weil der Thierkörper, entsprechend seiner viel mannichfaltigeren und vollkommneren Lebensthätigkeit, sich in viel mehr verschiedenen Richtungen differenziren und vervollkommnen konnte. Während wir daher das ganze Pflanzenreich in sechs Hauptklassen und achtzehn Klassen eintheilen konnten, müssen wir im Thierreich wenigstens sechszehn Hauptklassen und zwei und dreißig Klassen unterscheiden. Diese vertheilen sich in der Art, wie es die vorstehende systematische Uebersicht zeigt, auf die sechs verschiedenen Stämme des Thierreichs (S. 393).

Die Pflanzenthiere (Coelenterata), welche wir den übrigen fünf Stämmen des Thierreichs aus den angeführten Gründen gegenüberstellen, verdienen in mehr als einer Beziehung den Anfang zu machen. Denn abgesehen davon, daß dieselben in der That in ihrem gesammten Körperbau viel mehr von den übrigen fünf Stämmen verschieden sind, als diese unter sich, abgesehen ferner davon, daß auch ihre höchstentwickelten Formen nicht denjenigen Grad der Vollkommenheit und Differenzirung erreichen, wie die höchsten Formen der fünf anderen Stämme, schließen sich die Pflanzenthiere in mancher Hinsicht mehr den Pflanzen als den übrigen Thieren an. Insbesondere ist bei den fest gewachsenen Schwämmen und Korallen die äußere Körperform, der Mangel freier Ortsbewegung, die Stockbildung und die Fortpflanzung so ähnlich den entsprechenden Verhältnissen bei den Pflanzen, daß man dieselben noch im Beginn des vorigen Jahrhunderts ganz allgmein für wirkliche Pflanzen hielt. Der alte Name Zoophyta, was wörtlich übersetzt "Pflanzenthiere" bedeutet, war daher gar nicht übel gewählt. Die Bezeichnung Coelenterata erhielten dieselben von Leuckart, welcher 1848 zuerst ihre eigenthümliche Organisation erkannte und sie als eine ganz selbstständige Hauptabtheilung des Thierreichs aufstellte.
Durch die Bezeichnung Coelenterata wird der besondere anatomische Charakter ausgedrückt, durch welchen sich die Pflanzenthiere von allen übrigen Thieren unterscheiden. Bei den letzteren werden nämlich allgmein (nur die niedrigsten Formen ausgenommen) die vier verschiedenen Functionen der Ernährungsthätigkeit: Verdauung, Blutumlauf, Athmung und Ausscheidung durch vier ganz verschiedene Organsysteme bewerkstelligt, durch den Darm, das Blutgefäßsystem, die Athmungsorgane und die Harnapparate. Bei den Coelenteraten dagegen sind diese Functionen und ihre Organe noch nicht getrennt, und sie werden sämmtlich durch ein einziges System von Ernährungskanälen vertreten, durch das sogenannte Gastrovascularsystem oder den coelenterischen Darmgefäßapparat. Der Mund, der zugleich der After ist, führt in einen Magen, in welchen die übrigen Hohlräume des Körpers offen einmünden. Alle Pflanzenthiere leben im Wasser, und die allermeisten im Meere. Nur sehr wenige leben im süßen Wasser, nämlich die Süßwasserschwämme (Spongilla) und einige Urpolypen (Hydra, Cordylophora).
Der Stamm der Pflanzenthiere zerfällt in zwei verschiedene Hauptklassen, in die Schwämme oder Spongien und in die Nesselthiere oder Akalephen. Die letztere ist viel formenreicher und höher organisirt, als die erstere, welche die niederen Pflanzenthiere und darunter die ursprünglichen Stammformen des ganzen Stammes enthält. Bei den Schwämmen sind allgemein die ganze Körperform sowohl als die einzelnen Organe viel weniger differenzirt und vervollkommnet als bei den Nesselthieren. Insbesondere fehlen den Schwämmen allgemein die charakteristischen Nesselorgane, welche sämmtliche Nesselthiere besitzen. Das sind kleine, mit Gift gefüllte Bläschen, welche in großer Anzahl, meist zu vielen Millionen, in der Haut der Nesselthiere vertheilt sind, und bei Berührung derselben hervortreten und ihren Inhalt entleeren. Kleinere Thiere werden dadurch getötet; bei größeren bringt das Nesselgift, ganz ähnlich dem Gift unserer Brennnesseln, eine leichte Entzündung in der Haut hervor. Diejenigen von Ihnen, welche öfter in der See gebadet haben, werden dabei wohl schon bisweilen mit größeren Schirmquallen in Berührung gekommen sein und das unangenehme Gefühl kennen gelernt haben, das die Nesselorgane derselben hervorbringen. Bei den prachtvollen blauen Seeblasen oder Physalien wirkt das Gift so heftig, daß es den Tod des Menschen zur Folge haben kann.
Die Hauptklasse der Schwämme (Spongiae oder Porifera genannt), welche gewöhnlich als eine einzige Klasse aufgefaßt wird, kann man in zwei Gruppen oder Unterklassen vertheilen, in die Weichschwämme und Hartschwämme. Die Weichschwämme (Malacospongiae) besitzen gar keine harten Theile, kein Skelet, und ihr ganzer Körper besteht entweder aus einfachem ungesonderten Urschleim, oder aus nackten, amöbenartigen Urzellen. Wir unterscheiden in dieser Klasse zwei Ordnungen: die Urschwämme und die Schleimschwämme. Unter den Urschwämmen (Archispongiae) verstehen wir die längst ausgestorbenen hypothetischen Stammformen, aus denen sich die ganze Schwammklasse und somit auch der ganze Stamm der Cölenteraten entwickelt hat.
Es würden hierher gehören
1) die durch Urzeugung entstandenen Moneren, welche in ältester antelaurentischer Zeit dem ganzen Stamm den Ursprung gaben;
2) diejenigen Amöben oder einfachen nackten beweglichen Urzellen, welche aus diesen Moneren dadurch entstanden, daß sich im Inneren ein Kern von dem umgebenden Zellstoff differenzirte;
3) endlich die einfachsten vielzelligen Schwämme, welche sich aus den letzteren durch Coloniebildung entwickelten, d. h. dadurch, daß mehrere nackte Amoeben sich vereinigten und einen schleimigen Urschwammkörper darstellten (Prospongia).
Dadurch würden wir bereits unmittelbar zu der zweiten Ordnung geführt werden, den Schleimschwämmen (Myxospongiae), von denen noch heutzutage Halisarca Dujardinii in der Nordsee lebt. Das ist ein formloser Schleimkörper, welcher auf dem Thallus der Riementange oder Laminarien festsitzend angetroffen wird. Er besteht einzig und allein aus einer Gesellschaft von gleichartigen, nackten, amöbenähnlichen Zellen, welche in der Weise vereinigt sind, daß der Gesammtkörper von einem sehr unvollkommenen Canalsystem durchzogen wird. Diese Schleimschwämme, welche eigentlich nichts weiter als coelenterische Amöbengemeinden sind, verhalten sich zu den höchst differenzirten Nesselthieren ähnlich, wie die Stämme der Australneger, die noch keine Arbeitstheilung kenen, zu den höchstorganisirten Culturstaaten.
Die zweite Hauptabtheilung der Schwämme, die Hartschwämme (Sceletospongiae), haben sich offenbar erst später aus den Schleimschwämmen entwickelt. Sie unterscheiden sich von diesen dadurch, daß die nackten Amöben, welche den Weichkörper des Schwammes zusammensetzen, ein Hartgebilde oder Skelet ausscheiden, das dem ersteren als formgebende innere Stütze dient. Ja nach der verschiedenen chemischen Beschaffenheit dieses Skelets unterscheiden wir unter den Hartschwämmen vier Ordnungen: die Hornschwämme, Kieselschwämme, Kalkschwämme und Becherschwämme. Bei den Hornschwämmen (Ceratospongiae) besteht das Skelet bloß aus einer organischen Substanz, aus einer stickstoffhaltigen Kohlenstoffverbindung, welche Ihnen Allen als das faserige Maschengewebe des gewöhnlichen Badeschwammes (Euspongia officinalis) bekannt ist.
Dieses hornähnliche Fasergerüst, mit welchem wir uns jeden Morgen waschen, ist das eigentliche Skelet des Badeschwamms; alle seine Lücken sind im Leben ausgekleidet und die ganze Masse überzogen von dem schleimigen Weichkörper, der aus lauter Amöben zusammengesetzt ist. Aus diesen Hornschwämmen, die zunächst von den Schleimschwämmen abstammen, haben sich wahrscheinlich späterhin als drei divergente Zweige die drei übrigen Ordnungen, Kieselschwämme, Kalkschwämme und Becherschwämme entwickelt. Bei den Kieselschwämmen (Silicispongiae), zu denen auch unsere Süßwasserschwämme (Spongilla) gehören, besteht das Skelet aus vielen einzelnen Kieselnadeln, bei den Kalkschwämmen (Calcispongiae) dagegen aus Kalknadeln. Bei den Becherschwämmen (Petrospongiae) welche schon längst ausgestorben sind, aber massenhaft versteinert in den paläolitischen und besonders in den mesolithischen Schichten vorkommen, bildete das Skelet ein sehr regelmäßiges Gerüst von der Gestalt eines Bechers, eines Trichters, oder auch eines Hutpilzes.
Die Nesselthiere (Acelephae), welche sich durch die höhere Differenzirung der Organe und Gewebe und ganz besonders durch den Besitz der Nesselorgane von den Schwämmen unterscheiden, haben sich wahrscheinlich schon frühzeitig in der Primordialzeit aus diesen entwickelt. Man theilt diese Hauptklasse allgemein in drei Klassen, in die Korallen, Schirmquallen und Kammquallen (Vergl. Gen. Morph. II, Taf. III, S. L-LXI).
Die Klasse der Korallen (Corallia), wegen der Blumengestalt der einzelnen Individuen auch Blumenthiere (Anthozoa) genannt, schließt sich in vielfacher Beziehung auf das engste an die Schwämme an, as denen sie sich vielleicht unmittelbar entwickelt hat. Einige Kieselschwämme (z. B. Axinella polypoides) scheinen noch heutzutage unmittelbar den Uebergang zwischen beiden Klassen zu vermitteln. Die Gegenstücke oder Antimeren, d.h. die gleichartigen Hauptabschnitte des Körpers, welche strahlenförmig vertheilt, um die mittlere Hauptaxe des Körpers herumstehen, und deren Zahl bei den Schwämmen (wenn diese hier überhaupt differenzirt sind) schwankend ist, erscheinen bei den Korallen in verschiedener, aber sehr constanter Zahl. Je nach dieser Zahl unterscheiden wir unter den Korallen drei verschiedene Ordnungen, welche als drei Aeste einer gemeinsamen Stammform aufzufassen sind. Diese drei Ordnungen, deren Individuen oder Polypen aus je vier, sechs oder acht Gegenstücken regelmäßig zusammengesetzt erscheinen, sind die vierzähligen (Tetracorallia), die sechszähligen (Hexacorallia) und die achtzähligen Korallen (Octocorallia). {Abbildung Seite 400}

Die zweite Klasse der Nesselthiere bilden die Schirmquallen (Medusae) oder Polypenquallen (Hydremedusae). Während die Korallen meistens pflanzenähnliche Stöcke bilden, die auf dem Meeresboden festsitzen, schwimmen die Schirmquallen meistens in Form gallertiger Glocken frei im Meere umher. Jedoch giebt es auch unter ihnen zahlreiche, namentlich niedere Formen, welche auf dem Meeresboden festgewachsen sind und zierlichen Bäumchen gleichen. Die niedersten und einfachsten Angehörigen dieser Klasse die die bekannten Süßwasserpolypen (Hydra), welche bald grün, bald orangeroth, braun oder grau gefärbt sind. Gewöhnlich findet man sie in unsren Teichen an der Unterseite von Wasserlinsen ansitzen, als länglichrunde schleimige Körperchen von einer oder wenigen Linien Länge, die an dem freien Ende einen Mund und rings um diesen herum einen Kranz von 6-8 Fangarmen tragen.
Wir können sie als die wenig veränderten Nachkommen jener uralten Urpolypen (Archydrae) ansehen, welche während der Primordialzeit der ganzen Klasse der Hydromedusen und vielleicht der ganzen Hauptklasse der Nesselthiere den Ursprung gaben. Direkt oder indirekt können sich solche Hydrapolypen oder Hydroiden aus Weichschwämmen entwickelt haben. Von der Hydra kaum zu trennen sind diejenigen festsitzenden Hydroidpolypen (Campanularia, Sertularia, Tubularia), welche durch Knospenbildung frei schwimmende Medusen erzeugen, aus deren Eiern wiederum festsitzende Polypen entstehen. Diese frei schwimmenden Schirmquallen, welche in die drei Ordnungen der Zartquallen, Starrquallen und Scheibenquallen eingetheilt werden, haben meistens die Form eines Hutpilzes oder eines Regenschirms, von dessen Rand viele zarte und lange Fangfäden herabhängen. Sie gehören zu den schönsten und interessantesten Bewohnern des Meeres. Ihre merkwürdige Lebensgeschichte aber, insbesondere der verwickelte Generationswechsel der Polypen und Medusen, und die weitgehende Arbeitstheilung der Individuen, gehört zu den stärksten Zeugnissen für Wahrheit der Abstammungslehre.
Aus einem Zweige der Schirmquallen hat sich wahrscheinlich die dritte Klasse der Nesselthiere, die eigenthümliche Abtheilung der Kammquallen (Ctenophora) entwickelt. Diese Quallen, welche oft auch Rippenquallen oder Gurkenquallen genannt werden, besitzen einen gurkenförmigen Körper, welcher, gleich dem Körper der meisten Schirmquallen, krystallhell und durchsichtig wie geschliffenes Glas ist. Ausgezeichnet sind die Kammquallen oder Rippenquallen durch ihre eigenthümlichen Bewegungsorgane, nämlich acht Reihen von rudernden Wimperblättchen, die wie acht Rippen von einem Ende der Längsaxe (vom Munde) zum entgegengesetzten Ende verlaufen. Von den beiden Hauptabtheilungen derselben haben sich die Engmündigen (Stenostoma) wohl erst später aus den Weitmündigen (Eurystoma) entwickelt. Diese letzteren stammen wahrscheinlich direkt von Schirmquallen ab.
Indem wir nun den Stamm der Pflanzenthiere verlassen, wenden wir uns zu demjenigen Stamme des Thierreichs, welcher in genealogischer Beziehung die meisten Schwierigkeiten darbietet. Das ist das Phylum der Würmer (Vermes oder Helminthes). Wie schon vorher bemerkt, sind diese Schwierigkeiten höchst wahrscheinlich zum größten Theil dadurch bedingt, daß dieser Stamm die gemeinsame Ausgangsgruppe des ganzen Thierreichs ist, und daß er eine Masse von divergenten Aesten enthält, die sich theils zu ganz selbstständigen Würmerklassen entwickelt, theils aber in die ursprünglichen Wurzelformen der übrigen Stamme des Thierreichs umgebildet haben. Jeden der fünf übrigen Stämme konnten wir uns bildlich als einen hochstämmigen Baum vorstellen, dessen Stamm uns in seiner Verzweigung die verschiedenen Klassen, Ordnungen, Familien u. s. w. repräsentirt. Das Phylum der Würmer dagegen können wir nicht in einem solchen Bilde darstellen. Vielmehr würden wir uns dasselbe als einen niedrigen Busch oder Strauch zu denken haben, aus dessen Wurzel eine Masse von selbstständigen Zweigen nach verschiedenen Richtungen hin emporschießen. Und wenn man annimmt, daß das ganze Thierreich in dem Würmerstamm seine gemeinsame Wurzel hat, so würden die fünf übrigen Phylen als fünf einzelne Bäume zu denken sein, die aus jenem dichten Busche sich erheben, nur an der Wurzel unter einander und mit den zahlreichen Wurzelschößlingen (den Wurmklassen) zusammenhängend.
Die außerordentlichen Schwierigkeiten, welche die Systematik der Würmer schon aus diesem Grunde darbietet, werden nun aber dadurch noch sehr gesteigert, daß wir fast gar keine versteinerten Rest von ihnen besitzen. Die allermeisten Würmer besaßen und besitzen noch heute einen so weichen Leib, daß sie keine Spuren in den neptunischen Erschichten hinterlassen konnten. Auch die wenigen fossilen Reste von härteren Theilen, die wir von einigen Würmern besitzen, sind meistens so wenig charakteristisch, daß sie wenig mehr als die vormalige Existenz von jetzt ausgestorbenen Würmern anzeigen. Wir sind daher auch hier wieder vorzugsweise auf die Schöpfungsurkunden der Ontogenie und der vergleichenden Anatomie angewiesen, wenn wir den äußerst schwierigen Versuch unternehmen wollen, in das Dunkel des Würmerstammbaums einige hypothetische Streiflichter fallen zu lassen (Gen. Morph. II, Taf V, S. LXXVII-LXXXV).
Die zahlreichen Klassen, welche man im Stamme der Würmer unterscheiden kann, und welche jeder Zoologie in anderer Weise nach seinen subjektiven Anschauungen gruppirt, werden vielleicht am besten dadurch übersichtlich, daß man dieselben auf vier verschiedene Hauptklassen vertheilt. Diese wollen wir als Urwürmer, Weichwürmer, Sackwürmer und Gliedwürmer bezeichnet. Die Urwürmer enthalten, falls unsere einstämmige Descendenzhypothese richtig ist, jedenfalls die gemeinsamen Wurzelformen der übrigen Würmer, und wahrscheinlich des ganzen Thierreichs. Die Weichwürmer würden zum größten Theil selbstständige Wurmgruppen umfassen, die sich nicht zu höheren Thierstämmen entwickelt haben. Dagegen würden zu den Sackwürmern die Stammformen der Weichthiere und Wirbelthiere, zu den Gliedwürmern die Stammformen der Sternthiere und Gliedfüßer gehören. Die vier Hauptklassen der Würmer kann man in nachstehende 22 Ordnungen eintheilen. {Abbildung Seite 404...}

In der Hauptklasse der Urwürmer (Archelminthes) vereinigen wir diejenigen Thiere, welche jetzt gewöhnlich Infusionsthiere (Infusoria) im engeren Sinne genannt werden, mit denjenigen niedersten Wurzelformen des Stammes, aus denen sich die letzteren erst entwickelt haben können. Diese hypothetischen Wurzelformen würden wir den eigentlichen Infusorien (Ciliaten und Acineten) unter dem Namen der Urinfusorien oder Urahnthiere (Archezoa) gegenüberstellen können.
Als solche Archezoen, die also möglicherweise die ältesten gemeinsamen Ursprungsformen des ganzen Thierreichs sind, wären zu betrachten:
1) die durch Urzeugung entstandenen Moneren, welche in der ältesten antelaurentischen Zeit den Grund zum Thierreich, und zunächst zum Würmerstamm legten;
2) diejenigen Amöben, d. h. diejenigen ganz einfachen, nackten , beweglichen Urzellen, die sich aus jenen Moneren durch Differenzirung des centralen Kerns und des peripherischen Plasma entwickelten;
3) die einfachsten vielzelligen Würmer, welche dadurch entstanden, daß mehrere von jenen Amöben sich zur Bildung einer Colonie vereinigten, und nun durch Arbeitstheilung weiter entwickelten. An diese letzteren würden sich die echten Infusorien unmittelbar anschließen.
Möglicherweise leben noch heutzutage einige niederste Organismen, welche wahre Archezoen sind, nämlich gewisse Amoeben und die schmarotzenden Gregarinen. Vorsichtiger ist es aber jedenfalls, diese vorläufig als Protisten anzusehen, da uns ihre Abstammung unbekannt ist.
Als Infusionsthiere (Infusoria) im engeren Sinne werden heutzutage gewöhnlich nur die beiden Abtheilungen der Wimperinfusoria (Ciliata) und der Starrinfusorien (Acinetae) bezeichnet. Die meisten hierher gehörigen Thiere sind so klein, daß man sie mit dem bloßen Auge nicht sehen, und erst mit Hülfe starker Vergrößerungen ihre eigentliche Organisation erkennen kann. Gleich den meisten Protisten ersetzen sie aber durch Masse der Individuen, was ihnen an Körpergröße abgeht, und bevölkern das Meer und die süßen Gewässer in erstaunlichen Mengen. Vorzüglich gilt das von den Wimperinfusorien, welche die Hauptmasse der heutigen Infusionsthiere bilden. Ihren Namen führt diese ganze Gruppe von dem charakteristischen Wimperkleid, welches den ganzen Körper oder einen Theil desselben bedeckt, und mittelst dessen sie sich lebhaft umherbewegen.
Die Starrinfusorien dagegen sind wimperlos und sitzen unbeweglich fest; nur in frühester Jugend schwimmen sie mittelst eines vergänglichen Wimperkleides frei umher und sind dann von den Wimperthieren nicht zu unterscheiden. Unter den Wimperthieren schließen sich einige Formen unmittelbar an die frühesten Jugendzustände der Pflanzenthiere, andere an diejenigen der Würmer, der Sternthiere und der Weichthiere an. Einige Wimperthiere bilden den Uebergang zu den Strudelwürmern, anderen zu den Räderthieren, noch andere zu verschiedenen anderen Würmergruppen. In allen diesen Verhältnissen zusammengenommen finden wir genügend Grund, die bewimperten Infusorien (natürlich nicht die jetzt lebenden, sondern längst ausgestorbene Formen) als diejenigen Urwürmer zu betrachten, aus denen sich die übrigen Thierstämme direct oder indirect entwickelt haben.
Zunächst an die Urwürmer schließt sich von den übrigen Würmern die zweite Hauptklasse an, die Weichwürmer (Scolecida). Wir verstehen darunter die beiden tiefstehenden Klassen der Plattwürmer oder Platyelminthen und der Rundwürmer oder Nematelminthen. Die Klasse der Plattwürmer (Platyelminthes) führt ihren Namen von der blattförmigen Körpergestalt, die vom Rücken nach der Bauchseite stark zusammengedrückt ist. Die wahrscheinlichen Stammformen der ganzen Klasse sind die Strudelwürmer (Turbellaria), welche sich sowohl durch ihr Wimperkleid als durch ihre innere Organisation unmittelbar an die bewimperten Urwürmer oder Ciliaten anschließen.
Aus den frei im Wasser lebenden Strudelwürmern sind durch Anpassung an parasitische Lebensweise die schmarotzenden Saugwürmer (Trematoda) entstanden, und aus diesen durch weiter gehenden Parasitismus die Bandwürmer (Cestoda). Andrerseits haben sich vielleicht aus den Saugwürmern die Egel (Hirudinea) entwickelt, zu denen unser gewöhnlicher Blutegel gehört. Diesen vielleicht verwandt sind die Krallenwürmer (Onychophora). Als ein besonderer Zweig ist aus den Strudelwürmern die nahverwandte Gruppe der langen Schnurwürmer (Nemertina) hervorgegangen, welche größtentheils im Meere leben und wahrscheinlich die Stammeltern der Ringelwürmer sind.
Die Rundwürmer (Nematelminthes), die zweite Klasse der Weichwürmer, unterscheidet sich von der ersten Klasse, den Plattwürmern, durch ihre drehrunde oder cylindrische, nicht plattgedrückte Körpergestalt. Gleich vielen Plattwürmern sind auch die meisten Rundwürmer Schmarotzer, welche im Inneren anderer Thiere parasitisch leben. Frei im Meere lebend findet sich die eigenthümliche Gruppe der Pfeilwürmer (Chaetognathi oder Sagittae). Aus Rundwürmern, welche diesen wahrscheinlich sehr nahe standen, haben sich durch Anpassung an parasitische Lebensweise die Fadenwürmer (Nematoda) entwickelt, zu denen unter anderen die gemeinen Spulwürmer, die berühmten Trichinen, Medinawürmer und die viele anderen Schmarotzer des Menschen gehören. Noch weiter entartete Parasiten dieser Klasse sind die mit einem Hakenrüssel versehenen Kratzwürmer (Acanthocephala oder Echinorhynchi). Wahrscheinlich ist die gemeinsame Stammform aller dieser Rundwürmer ein unbekannter Wurm, welcher sich aus einem Zweige der Plattwürmer entwickelt hat.
Eine ganz eigenthümlich und sehr merkwürdige Astgruppe des Würmerstammes bildet die dritte Hauptklasse, die Sackwürmer (Himatega). Wir fassen unter dieser Bezeichnung die beiden Klassen der Mosthiere oder Bryozoen und der Mantelthiere oder Tunikaten zusammen. Bisher stellte man diese beiden Thierklassen im zoologischen Systeme gewöhnlich zu dem Stamme der Weichthiere oder Mollusken und setzte sie hier den echten Weichthieren (Muscheln, Schnecken u.s. w.) als Weichthierartige (Molluscoidea) gegenüber. Diese Auffassung läßt sich dadurch rechtfertigen, daß allerdings die echten Weichthiere wahrscheinlich von denselben abstammen, und zwar von den Mosthieren. Allein andrerseits erscheinen die Mantelthiere näher mit den Wirbelthieren verwandt, und aus diesem Grunde dürfte es wohl das Beste sein, beide Klassen wieder in die vielgestaltige Würmergruppe zurückzustellen, und als verbindende Zwischenformen zwischen den niederen Würmern einerseits und den Mollusken und Wirbelthieren andrerseits aufzufassen.
So wenig es passend sein würde, die Mantelthiere auf Grund ihrer offenbaren Blutsverwandtschaft mit den Wirbelthieren gradezu im System zu vereinigen, so wenig vortheilhaft ist es auch für die systematische Auffassung, wenn man die Mosthiere mit den echten Weichthieren vereinigt. Wie die meisten Klassen der Sackwürmer übrigens eigentlich untereinander mit den niederen Würmern zusammenhängen, ist uns heutzutage noch sehr unklar, obwohl an ihrer Abstammung von niederen Würmern (entweder von Weichwürmern oder direct von Urwürmern) nicht zu zweifeln ist.
Die Klasse der Mosthiere (Bryozoa) enthält sehr kleine, zierliche Würmer, welche in Form mosähnlicher Bäumchen oder Polster auf Steinen und anderen Gegenständen im Meere (selten im süßen Wasser ) festsitzen. Früher wurden dieselben gewöhnlich zu den Pflanzenthieren gerechnet, und in der That sind sie manchen von diesen sehr ähnlich. Insbesondere gleichen sie den Hydroidpolypen durch ihre äußere Form, durch einen Fühlerkranz, welcher den Mund umgiebt, und durch die Art und Weise, in welcher zahlreiche Individuen zu baumförmigen und rindenförmigen Colonien vereinigt leben. Allein durch ihre innere Organisation sind die Mosthiere ganz von den Pflanzenthieren verschieden und schließen sich vielmehr einerseits den niederen Würmern, andrerseits den niedersten Weichthieren, den Spiralkiemern oder Spirobranchien an. Namentlich sind die Jugendformen der letzteren den Mosthieren sehr ähnlich, und hierauf vorzüglich, sowie auch auf ihre anatomische Verwandtschaft gründet die Vermuthung, daß die Mosthiere nächste Verwandte derjenigen ausgestorbenen Würmer sind, aus denen sich der Stamm der Mollusken, und zwar zunächst die Armkiemer, entwickelten. Von den beiden Hauptabtheilungen der Mosthiere stehen die höheren, diejenigen mit einem Kragen (Phylactolaema), den Armkiemern näher, als die niederen Mosthiere, ohne Kragen (Gymnolaema).
In ganz ähnlicher Beziehung wie die Mosthiere zu den Weichthieren, steht die zweite Klasse der Sackwürmer, die Mantelthiere (Tunicata), zu den Wirbelthieren. Diese höchst merkwürdige Thierklasse lebt im Meere, wo die einen (die Seescheiden oder Chthonascidien) auf dem Boden festsitzen, die anderen (die Seetonnen oder Nektascidien) frei umherschwimmen. Bei allen besitzt der ungegliederte Körper die Gestalt eines einfachen tonnenförmigen Sackes, welcher von einem dicken knorpelähnlichen Mantel eng umschlossen ist. Dieser Mantel besteht aus derselben stickstofflosen Kohlenstoffverbindung, welche im Pflanzenreich als "Cellulose" eine so große Rolle spielt und den größten Theil der pflanzlichen Zellmembranen und somit auch des Holzes bildet. Gewöhnlich besitzt der tonnenförmige Körper keinerlei äußere Anhänge. Niemand würde darin irgend eine Spur von Verwandtschaft mit den hoch differenzirten Wirbelthiren erkennen.
Und doch kann diese nicht mehr zweifelhaft sein, seitdem vor zwei Jahren die Untersuchungen von Kowalewsky plötzlich darüber ein höchst überraschendes und merkwürdiges Licht verbreitet haben. Aus diesen hat sich nämlich ergeben, daß die individuelle Entwickelung der festsitzenden einfachen Seescheiden (Ascidia, Phallusia) in den wichtigsten Beziehungen mit derjenigen des niedersten Wirbelthieres, des Lanzetthieres (Amphioxus lanceolatus) übereinstimmt. Insbesondere besitzen die Jugendzustände der Ascidien die Anlage des Rückenmarks und des darunter gelegenen Rückenstrangs (Chorda dorsalis) d. h. die beiden wichtigsten und am meisten charakteristischen Organe des Wirbelthierkörpers. Unter allen uns bekannten wirbellosen Thieren besitzen demnach die Mantelthiere zweifelsohne die nächste Blutsverwandtschaft mit den Wirbelthieren, und sind als nächste Verwandte derjenigen Würmer zu betrachten, aus denen sich dieser letztere Stamm entwickelt hat.
Die vierte und letzte Hauptklasse des Würmerstammes, die der Gliedwürmer (Colelminthes) zeichnet sich vor den drei übrigen Klassen durch die deutliche Gliederung des Körpers aus, d. h. durch die Zusammensetzung desselben aus mehreren, in der Längsaxe hinter einander gelegenen Abschnitten, den Gliedern, Segmenten oder Folgestücken (Metameren). Wir unterscheiden in dieser Hauptklasse die drei Klassen der Sternwürmer, Ringelwürmer und Räderthiere.
Die Sternwürmer (Gephyrea) sind langgestreckte, drehrunde oder walzenförmige Würmer, bei denen die Körpergliederung äußerlich wenigstens, erst sehr undeutlich ausgesprochen ist. Sie leben alle auf dem Boden des Meeres, entweder im Sand oder Schlamm vergraben, oder in Löchern, welche sie in die Felsen bohren. An sich sind die Sternwürmer von keinem besonderen Interesse, wohl aber dadurch, daß sie wahrscheinlich die nächsten Verwandten der Panzerwürmer oder Phraktelminthen sind, d. h. derjenigen gegliederten Würmer, aus denen sich der Stamm der Echinodermen entwickelt hat.
Die zweite Klasse der Gliedwürmer bildet die umfangreiche Abtheilung der Ringelwürmer (Annelida). Dahin gehören einerseits die nackten Regenwürmer und ihre Verwandten, welche wir als Kahlwürmer (Drilomorpha) zusammenfassen, andrerseits die mit Borsten bewaffneten Borstenwürmer (Chaetopoda), die im Meere frei umherkriechenden Raubwürmer (Vagantia), die in Röhren versteckten Röhrenwürmer (Tubicolae) und die frei schwimmenden Ruderwürmer (Gymnocopa). Endlich kann man als eine dritte Ordnung mit den Ringelwürmern auch die Bärwürmer (Arctisca) vereinigen, kleine im Mose, auf Baumrinden u. s. w. sehr häufige Würmer, welche wegen ihrer acht Beinstummel gewöhnlich (aber wohl mit Unrecht) zu den Spinnen gerechnt werden. Die meisten Ringelwürmer erreichen einen höheren Organisationsgrad als die übrigen Würmer, und entwickeln den eigentlichen Wurmtypus zu seiner höchsten Ausbildung. Viele schließen sich dadurch bereits unmittelbar an den Stamm der Gliedfüßer oder Arthopoden an, und es ist möglich, daß dieser wirklich von ausgestorbenen Ringelwürmern abstammt. Wahrscheinlicher jedoch ist es, daß er sich aus der dritten Klasse der Gliedwürmer, aus den Räderthieren entwickelt hat.
Die Räderthiere oder Räderwürmer (Rotatoria oder Rotifera) gehören zu denjenigen Klassen des Thierreichs, deren systematische Stellung den Zoologen von jeher die größten Schwierigkeiten bereitet hat. Meist sind es ganz kleine, nur durch das Mikroskop erkennbare Thierchen, welche mittelst eines besonderen, wimpernden Räderorgans im Wasser umherschwimmen; selten sitzen sie festgewachsen auf Wasserpflanzen und dergleichen auf. Einerseits schließen sie sich durch ihre niedersten Formen unmittelbar den Weichwürmern und zwar den Strudelwürmern (in mancher Beziehung auch den Bärwürmern) an. Andrerseits bilden sie in ihren höchst entwickelten Formen bereits den †bergang zu den Gliedfüßern (Arthropoda). Aller Wahrscheinlichkeit nach haben sich diese letzteren, und zwar zunächst krebsartige Thiere (Nauplius) aus Würmern entwickelt, welche von den heutigen Räderthieren im System kaum zu trennen waren.
Indem wir nun aus der buntgemischten Gesellschaft des vielgestaltigen Würmerstammes heraustreten, wollen wir nach einander noch kurz die vier höheren Stämme des Thierreichs betrachten, die sich aus verschiedenen Zweigen des ersteren entwickelt haben, die Weichthiere, Sternthiere, Gliedfüßer und Wirbelthiere. Unzweifelhaft der tiefststehende von diesen Stämmen, wenigstens in Bezug auf die morphologische Ausbildung, ist der Stamm der Weichthiere (Mollusca). Nirgends begegnen wir hier der charakteristischen Gliederung (Artikulation oder Metamerenbildung) des Körpers, welche schon die Gliedwürmer auszeichnete, und welche bei den übrigen drei Stämmen, den Sternthieren, Gliedfüßern und Wirbelthieren, die wesentlichste Ursache der höheren Formentwickelung, Differenzirung und Vervollkommnung wird. Vielmehr stellt bei allen Weichthieren, bei allen Muscheln, Schnecken u. s. w. der ganze Körper einen einfachen ungegliederten Sack dar, in dessen Höhle die Eingeweide liegen. Das Nervensystem besteht aus mehreren einzelnen (gewöhnlich drei), nur locker mit einander verbundenen Knotenpaaren, und nicht aus einem gegliederten Strang, wie bei den Sternthieren, Gliedfüßern und Wirbelthieren. Aus diesen und vielen anderen anatomischen Gründen halte ich den Weichthierstamm (trotz der höheren physiologischen Ausbildung seiner vollkommensten Formen) für den morphologisch niedersten unter den vier höheren Tierstämmen.
Wenn wir die Himategen oder Molluscoiden, die gewöhnlich mit dem Weichthierstamm vereinigt werden, aus den angeführten Gründen ausschließen, so behalten wir als echte Mollusken folgende vier Klassen: die Spiralkiemer, Blattkiemer, Schnecken und Pulpen. Die beiden niederen Mulluskenklassen, Spiralkiemer und Blattkiemer, besitzen weder Kopf noch Zähne, und man kann sie daher als Kopflose (Acephala) oder Zahnlose (Anodontoda) in einer Hauptklasse vereinigen. Diese Hauptklasse wird auch häufig als die der Muscheln (Conchifera) oder Zweiklappigen (Bivalva) bezeichnet, weil alle Mitglieder derselben eine zweiklappige Kalkschale besitzen. Den Muscheln oder Kopflosen gegenüber kann man die beiden höheren Weichthierklassen, Schnecken und Pulpen, als Kopfträger (Eucephala) oder Zahnträger (Odontophora) in einer zweiten Hauptklasse zusammenfassen, weil sowohl Kopf als Zähne bei ihnen ausgebildet sind.
Bei der großen Mehrzahl der Weichthiere ist der weiche sackförmige Körper von einer Kalkschale oder einem Kalkgehäuse geschützt, welches bei den Muscheln (sowohl Spiralkiemern als Blattkiemern) aus zwei Klappen, bei den Kopfträgern dagegen (Schnecken und Pulpen) aus einer meist gewundenen Röhre (dem sogenannten "Schneckenhaus") besteht. Trotzdem diese harten Skelete massenhaft in allen neptunischen Schichten sich versteinert finden, sagen uns dieselben dennoch nur sehr wenig über die geschichtliche Entwickelung des Stammes aus. Denn diese fällt größtentheils in die Primordialzeit.
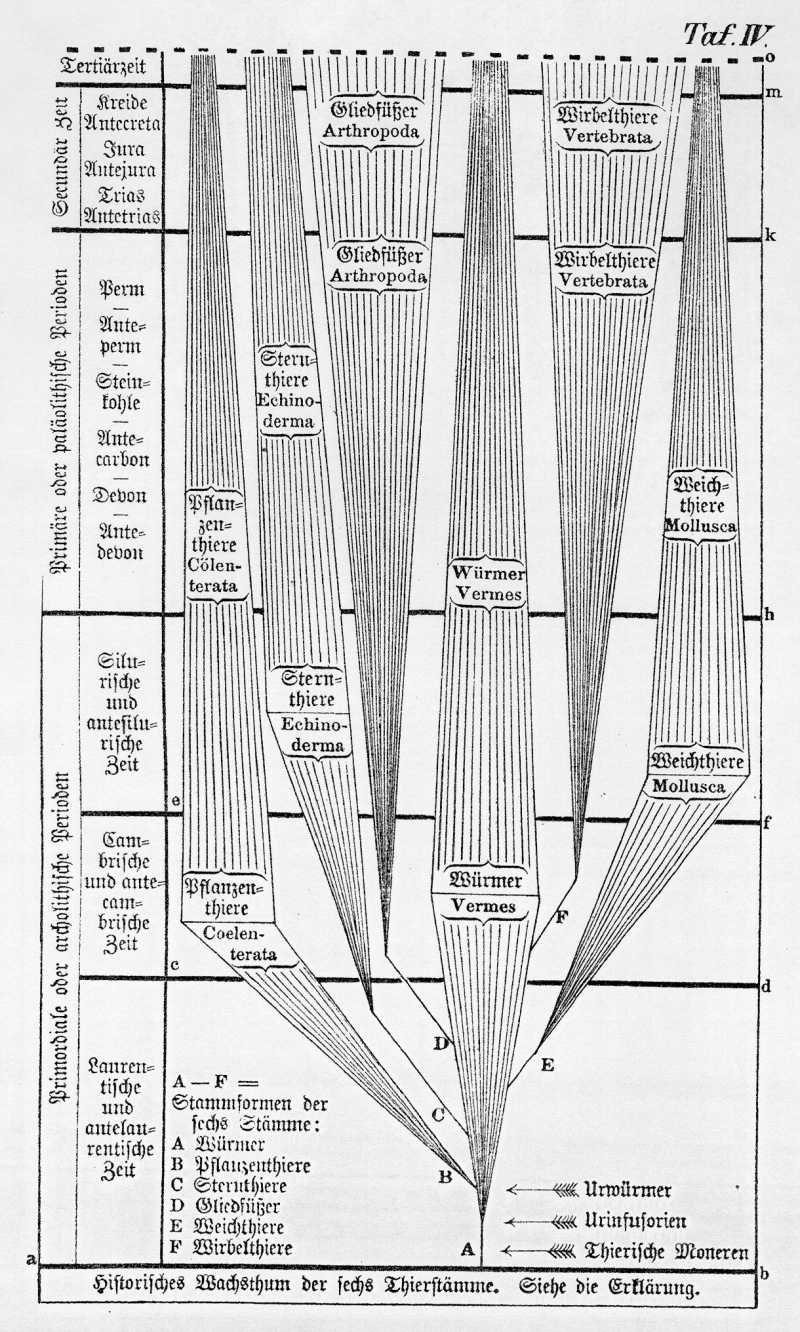
Selbst schon in den silurischen Schichten finden wir alle vier Klassen der Weichthiere neben einander versteinert vor, und dies beweist deutlich, in Uebereinstimmung mit vielen anderen Zeugnissen, daß der Weichthierstamm damals schon eine mächtige Ausbildung erreicht hatte, als die höheren Stämme, namentlich Gliedfüßer und Wirbelthiere, kaum über den Beginn ihrer historischen Entwickelung hinaus waren. In den darauf folgenden Zeitaltern, besonders zunächst im primären und weiterhin im secundären Zeitraum, dehnten sich diese höheren Stämme mehr und mehr auf Kosten der Mollusken und Würmer aus, welche ihnen im Kampfe um das Dasein nicht gewachsen waren, und dem entsprechend mehr und mehr abnahmen. Die jetzt noch lebenden Weichthiere und Würmer sind nur als ein verhältnißmäßig schwacher Rest von der mächtigen Fauna zu betrachten, welche in primordialer und primärer Zeit über die anderen Stämme ganz überwiegend herrschte. (Vergl. Taf. III und IV nebst Erklärung).
In keinem Thierstamm zeigt sich deutlicher, als in dem der Mollusken, wie verschieden der Werth ist, welchen die Versteinerungen für die Geologie und für die Phylogenie besitzen. Für die Geologie sind die verschiedenen Arten der versteinerten Weichthierschalen von der größten Bedeutung, weil dieselben als "Leitmuscheln" vortreffliche Dienste zur Charakteristik der verschiedenen Schichtengruppen und ihres relativen Alters leisten. Für die Genealogie der Mollusken dagegen besitzen sie nur sehr geringen Werth, weil sie einerseits Körpertheile von ganz untergeordneter morphologischer Bedeutung sind, und weil andererseits die eigentliche Entwickelung des Stammes in die ältere Primordialzeit fällt, aus welcher uns keine deutlichen Versteinerungen erhalten sind. Wenn wir daher den Stammbaum der Mollusken construiren wollen, so sind wir vorzugsweise auf die Urkunden der Ontogenie und der vergleichenden Anatomie angewiesen, aus denen sich etwa folgendes ergiebt. (Gen. Morph. II, Taf. VI, S. CII bis CXVI).
Von den vier uns bekannten Klassen der echten Weichthiere stehen auf der niedersten Stufe die in der Tiefe des Meeres festgewachsenen Spiralkiemer (Spirobranchia), oft auch unpassed als Armfüßer (Brachiopoda) bezeichnet. Von dieser Klasse leben gegenwärtig nur noch wenige Formen, einige Arten von Lingula, Terebratula und Verwandte; schwache Ueberbleibsel von der mächtigen und formenreichen Gruppe, welche die Spiralkiemer in älteren Zeiten der Erdgeschichte darstellten. In der Silurzeit bildeten sie die Hauptmasse des ganzen Weichthierstammes. Aus der Uebereinstimmung ihrer Jugendzustände mit denjenigen der Mosthiere schließen wir, daß sie sich aus dieser Klasse der Sackwürmer entwickelt haben.
Die zweite Weichthierklasse, die Blattkiemer (Elatobranchia oder Lamellibranchia) besitzen gleich den Spiralkiemern eine zweiklappige Schale. Es gehören hierher die meisten jetzt lebenden Muschelthiere des Meeres und die wenigen Muscheln unserer süßen Gewässer (Unio, Anodonta, Cyclas) Obwohl noch ohne Kopf und Gebiß, gleich den Spiralkiemern, sind sie doch im Uebrigen höher als diese organisirt und haben sich wahrscheinlich erst später aus einem Zweige jener Klasse entwickelt.
Von den kopftragenden Weichthieren stehen den kopflosen Muschen am nächsten die Schnecken (Cochlides oder Cephalophora), von denen wiederum die große Mehrzahl im Meere lebt, nur wenige im süßen Wasser, oder lutfathmend auf dem Lande. Durch die Stummelköpfe (Porocephala) sind die höher entwickelten Kopfschnecken (Delocophala) unmittelbar mit den Blattkiemern verbunden, von denen sie sich wahrscheinlich schon in früher Primordialzeit abgezweigt haben.
Die vierte und letzte, und zugleich die höchst entwickelte Klasse der Mollusken bilden die Pulpen, auch Tintenfische oder Kopffüßer genannt (Cephalopoda). Die Pulpen, welche noch jetzt in unseren Meeren leben, die Sepien, Kalmare, Argonautenboote und Perlboote, sind gleich den wenigen Spiralkiemern der Gegenwart nur dürftige Reste von der formenreichen Schaar, welche diese Klasse in den Meeren der primordialen, primären und secundären Zeit bildete. Die zahlreichen versteinerten Ammonshörner (Ammonites), Perlboote (Nautilida) und Donnerkeile (Belemnites) legen noch heutzutage von jenem längst erloschenen Glanze des Stammes Zeugniß ab. Wahrscheinlich haben sich die Pulpen aus einem niederen Zweige der Schneckenklasse, aus den Flügelschnecken (Pteropoden) oder Verwandten derselben entwickelt.
Die verschiedenen Unterklassen, Legionen und Ordnungen, welche man in den vier Molluskenklassen unterscheidet, und deren systematische Reihenfolge { Tabelle Seite 415 }

Ihnen die vorstehende Tabelle anführt, liefern in ihrer historischen und ihrer entsprechenden systematischen Entwickelung mannichfache Beweise für die Gültigkeit des Fortschrittsgesetzes. Da jedoch diese untergeordneten Molluskengruppen an sich weiter von keinem besonderen Interesse sind, verweise ich Sie auf den ausführlichen Stammbaum der Weichthiere, welchen ich in meiner generellen Morphologie gegeben habe, und wende mich sogleich weiter zur Betrachtung des Sternthierstammes.
Die Sternthiere (Echinodermata), zu welchen die vier Klassen der Seesterne, Seelilien, Seeigel und Seewalzen gehören, sind eine der interessantesten, und dennoch eine der wenigst bekannten Abtheilungen des Thierreichs. Jeder von Ihnen, der einmal an der See war, wird wenigsten zwei Formen derselben, die Seesterne und Seeigel gesehen haben. Wegen ihrer sehr eigenthümlichen Organisation sind die Sternthiere als ein ganz selbstständiger Stamm des Thierreichs zu betrachten, und namentlich gänzlich von den Pflanzenthieren oder Cölenteraten zu trennen, mit denen sie noch jetzt oft irrthümlich als Strahlthiere oder Radiaten zusammengefaßt werden (so z. B. von Agassiz, welcher auch diesen Irrthum Cuvier's neben manchen anderen noch heute vertheidigt). Eher als mit den Pflanzenthieren könnte man die Sternthiere mit den Würmern oder selbst mit den Gliedfüßern vereinigen.
Alle Echinodermen sind ausgezeichnet und zugleich von allen anderen Thieren verschieden durch einen sehr merkwürdigen Bewegungsapparat. Dieser besteht in einem verwickelten System von Canälen oder Röhren, die von außen mit Seewasser gefüllt werden. Das Seewasser wird in dieser Wasserleitung theils durch schlagende Wimperhaare, theils durch Zusammenziehungen der muskulösen Röhrenwände selbst, die Gummischläuchen vergleichbar sind, fortbewegt. Aus den Röhren wird das Wasser in sehr zahlreiche hohle Füßchen hinein gepreßt, welche dadurch prall ausgedehnt und nun zum Gehen und zum Ansaugen benutzt werden.
Außerdem sind die Sternthiere auch durch eine eigenthümliche Verkalkung der Haut ausgezeichnet welche bei den meisten zur Bildung eines festen, geschlossenen, aus vielen Platten zusammengesetzten Panzers führt. Bei fast allen Echinodermen ist der Körper aus fünf Strahltheilen (Gegenstücken oder Antimeren) zusammengesetzt, welche rings um die Hauptaxe des Körpers sternförmig herum stehen und sich in dieser Axe berühren. Nur bei einigen Seesternarten steigt die Zahl dieser Strahltheile über fünf hinaus, auf 6-9, 10-12, oder selbst 20- 40; und in diesem Falle ist die Zahl der Strahltheile bei den verschiedenen Individuen der Species meist nicht beständig, sondern wechselnd.
Die geschichtliche Entwickelung und der Stammbaum der Echinodermen werden uns durch ihre zahlreichen und meist vortrefflich erhaltenen Versteinerungen, durch ihre sehr merkwürdige individuelle Entwickelungsgeschichte und durch ihre interessante vergleichende Anatomie so vollständig enthüllt, wie es außerdem bei keinem anderen Thierstamme, selbst die Wirbelthiere nicht ausgenommen, der Fall ist. Durch eine kritische Benutzung jener drei Archive und eine denkende Vergleichung ihrer Resultate gelangen wir zu folgender Genealogie der Sternthiere, die ich in meiner generallen Morphologie begründet habe (Gen. Morph. II, Taf. IV. S. LXII.-LXXVII).
Die älteste und ursprünglichste Gruppe der Sternthiere, die Stammform des ganzen Phylum, ist die Klasse der Seesterne (Asterida). Dafür spricht außer zahlreichen und wichtigen Beweisgründen der Anatomie und Entwickelungsgeschichte vor allen die hier noch unbeständige und wechselnde Zahl der Strahltheile oder Antimeren, welche bei allen übrigen Echinodermen ausnahmslos auf fünf fixirt ist. Jeder Seestern besteht aus einer mittleren kleinen Körperscheibe, an deren Umkreis in einer Ebene fünf oder mehr lange gegliederte Arme befestigt sind. Jeder Arm des Seesterns entspricht in seiner ganzen Organisation wesentlich einem gegliederten Wurme aus der Hauptklasse der Gliedwürmer oder Colelminthen. Ich betrachte daher den Seestern als einen echten Stock oder Cormus von fünf oder mehr geliederten Würmern, welche mit dem einem Ende ihres Körpers verwachsen sind. Hier haben sie sich eine gemeinschaftliche Mundöffnung und eine gemeinsame Verdauungshöhle (Magen) gebildet, die in der mittleren Körperscheibe liegen. Das verwachsenen Ende, welches in die gemeinsame Mittelscheibe mündet, ist wahrscheinlich das Hinterende der ursprünglichen selbstständigen Würmer; denn das engegengesetzte freie Ende trägt zusammengesetzte Augen, wie sie nur noch an dem Kopfe der Gliedfüßer (Arthropoden) vorkommen.
In ganz ähnlicher Weise sind auch bei den ungegliederten Würmern bisweilen mehrere Individuen zur Bildung eines sternförmigen Stockes vereinigt. Das ist namentlich bei den Botrylliden der Fall, zusammengesetzte Seescheiden oder Ascidien, welche zur Klasse der Mantelthiere (Tunicaten) gehören. Auch hier sind die einzelnen Würmer mit ihrem hinteren Ende, wie ein Rattenkönig, verwachsen, und haben sich hier eine gemeinsame Auswurfsöffnung, eine Centralkloake gebildet, während am vorderen Ende noch jeder Wurm seine eigene Mundöffnung besitzt. Bei den Seesternen würde die letztere im Laufe der historichen Stockentwicklung zugewachsen sein, während sich die Centralkloake zu einem gemeinsamen Mund für den ganzen Stock ausbildete.
Die Seesterne würden demnach Würmerstöcke sein, welche sich entweder durch sternförmige Knospenbildung oder durch sternförmige Verwachsung aus echten gegliederten Würmern oder Colelminthen entwickelt haben. Diese Hypothese wird auf das Stärkste durch die vergleichende Anatomie und Ontogenie der gegliederten Seesterne (Colastra) und der gegliederten Würmer (Colelminthes) gestützt. Unter den letzteren stehen in Bezug auf den inneren Bau einerseits die Sternwürmer (Gephyrea), andrerseits die Ringelwürmer (Annelida) den einzelnen Armen oder Strahltheilen der Seesterne, d. h. den ursprünglichen Einzelwürmern, ganz nahe. Was aber das Wichtigste ist, aus den Eiern der Echinodermen entwickeln sich bewimperte Larven, welche nicht die geringste Aehnlichkeit mit den erwachsenen Sternthieren zeigen, dagegen den Larven gewisser Sternwürmer und mancher Ringelwürmer höchst ähnlich sind. Diese bilateral- symmetrischen Larven, welche keine Spur von der regulär- strahligen Sternform des erwAchsenen Echinoderms besitzen, erzeugen das letztere durch einen höchst merkwürdigen Generationswechsel, welcher in dieser Weise nur noch bei einigen Sternwürmern (Sipunculiden) und Schnurwürmern (Nemertinen) vorkömmt (Gen. Morph. II, 95-99).
Alle diese und viele andere Gründe legen das deutlichste Zeugniß für die Richtigkeit meiner Hypothese ab. Ich habe diese Stammhypothese 1866 aufgestellt, ohne eine Ahnung davon zu haben, daß auch noch versteinerte Gliedwürmer exitiren, welche vollkommen jenen hypothetisch vorausgesetzten Stammformen entsprechen.
Solche sind aber inzwischen wirklich bekannt geworden. In einer Abhandlung "über ein Aequivalent der takonischen Schiefer Nordamerikas in Deutschland" beschrieben 1867 Geinitz und Liebe eine Anzahl von gegliederten silurischen Würmern, welche vollkommen den von mir gemachten Voraussetzungen entsprechen.
Diese höchst merkwürdigen Würmer kommen in den Dachschiefern von Wurzbach im reussischen Oberlande zahlreich in vortrefflich erhaltenem Zustande vor. Sie haben ganz den Bau eines gegliederten Seesternarms, und müssen offenbar einen festen Hautpanzer, ein viel härteres und festeres Hautskelet besessen haben, als es sonst bei den Würmern vorkommt. Die Zahl der Körperglieder oder Metameren ist sehr beträchtlich, so daß die Würmer bei einer Breite von 1/4-1/2 Zoll eine Länge von 2-3 Fuß und mehr erreichten. Die vortrefflich erhaltenen Abdrücke, namentlich von Phyllodocites thuringiacus und Crossopodia Henrici, gleichen so sehr den skeletirten Armen mancher gegliederten Seesterne (Colastra), daß ich an ihrer wirklichen Blutsverwandtschaft kaum mehr zweifle. Ich bezeichne diese uralte Würmergruppe, zu welcher höchstwahrscheinlich die Stammväter der Seesterne gehört haben, als Panzerwürmer (Phractelminthes). Wahrscheinlich standen sie in ihrer Organisation zwischen Sternwürmern (Gephyreen) und Ringelwürmern (Anneliden) in der Mitte.
Aus der Klasse der Seesterne, welche die ursprüngliche Form des sternförmigen Wurmstockes am getreuesten erhalten hat, haben sich die { Tabelle Seite 420 } drei anderen Klassen der Echinodermen offenbar erst später entwickelt. Am wenigsten von ihnen entfernt haben sich die Seelilien (Crinoida), welche aber die freie Ortsbewegung der übrigen Sternthiere aufgegeben, sich festgesetzt, und dann einen mehr oder minder langen Stiel entwickelt haben. Die ursprünglichen Wurmindividuen sind zwar bei den Crinoiden nicht mehr so selbstständig und ausgebildet erhalten, wie bei den Seesternen; aber dennoch bilden sie stets mehr oder minder gegliederte, von der gemeinsamen Mittelscheibe abgesetzte Arme. Wir können daher die Seelilien mit den Seesternen zusammen in der Hauptklasse der Gliederarmigen (Colobrachia) vereinigen.

In den beiden anderen Echinodermenklassen, bei den Seeigeln und Seewalzen, sind die gegliederten Arme nicht mehr als selbstständige Körpertheile erkennbar, vielmehr durch weitgehende Centralisation des Stockes vollkommen in der Bildung der gemeinsamen, aufgeblasenen Mittelscheibe aufgegangen, so daß diese jetzt als eine einfache armlose Büchse oder Kapsel erscheint. Der ursprüngliche Individuenstock ist scheinbar dadurch wieder zum Formwerth eines einfachen Individuums, einer einzelnen Person, herabgesunken.
Wir können daher diese beiden Klassen als Armlose (Lipobrachia) den Gliederarmigen gegenübersetzen. Die erste Klasse derselben, die Seeigel (Echinida) führen Namen von den zahlreichen, oft sehr großen Stacheln, welche die feste, aus Kalkplatten sehr künstlich zusammengesetzte Schale bedecken. Die Schale selbst hat die Grundform einer fünfseitigen Pyramide. Wahrscheinlich haben sich die Seeigel unmittelbar aus einem Zweige der Seesterne, vielleicht im Zusammenhang mit einem Zweige der Seelilien entwickelt. Die einzelnen Abtheilungen der Seeigel bestätigen in ihrer historischen Aufeinanderfolge ebenso wie die Ordnungen der Seelilien und Seesterne, welche Ihnen die nebenstehende Tabelle aufführt, in ausgezeichneter Weise die Gesetze des Fortschritts und der Differenzirung. In jeder jüngeren Periode der Erdgeschichte sehen wir die einzelnen Klassen an Mannichfaltigkeit und Vollkommenheit zunehmen (Gen. Morph. II, Taf. IV).
Während uns die Geschichte dieser drei Sternthierklassen durch die zahlreichen und vortrefflich erhaltenen Versteinerungen sehr genau erzählt wird, wissen wir dagegen von der geschichtlichen Entwickelung der vierten Klasse, der Seewalzen (Holothuriae), fast Nichts. Die Skeletbildung der Haut ist hier sehr unvollkommen und daher konnten keine deutlichen Reste von ihrem langgestreckten walzenförmigen wurmähnlichen Körper in fossilem Zustande erhalten bleiben. Dagegen läßt sich aus der vergleichenden Anatomie der Holothurien erschließen, daß dieselben wahrscheinlich aus einer Abtheilung der Seeigel durch Erweichung des Hautskelets entstanden sind.
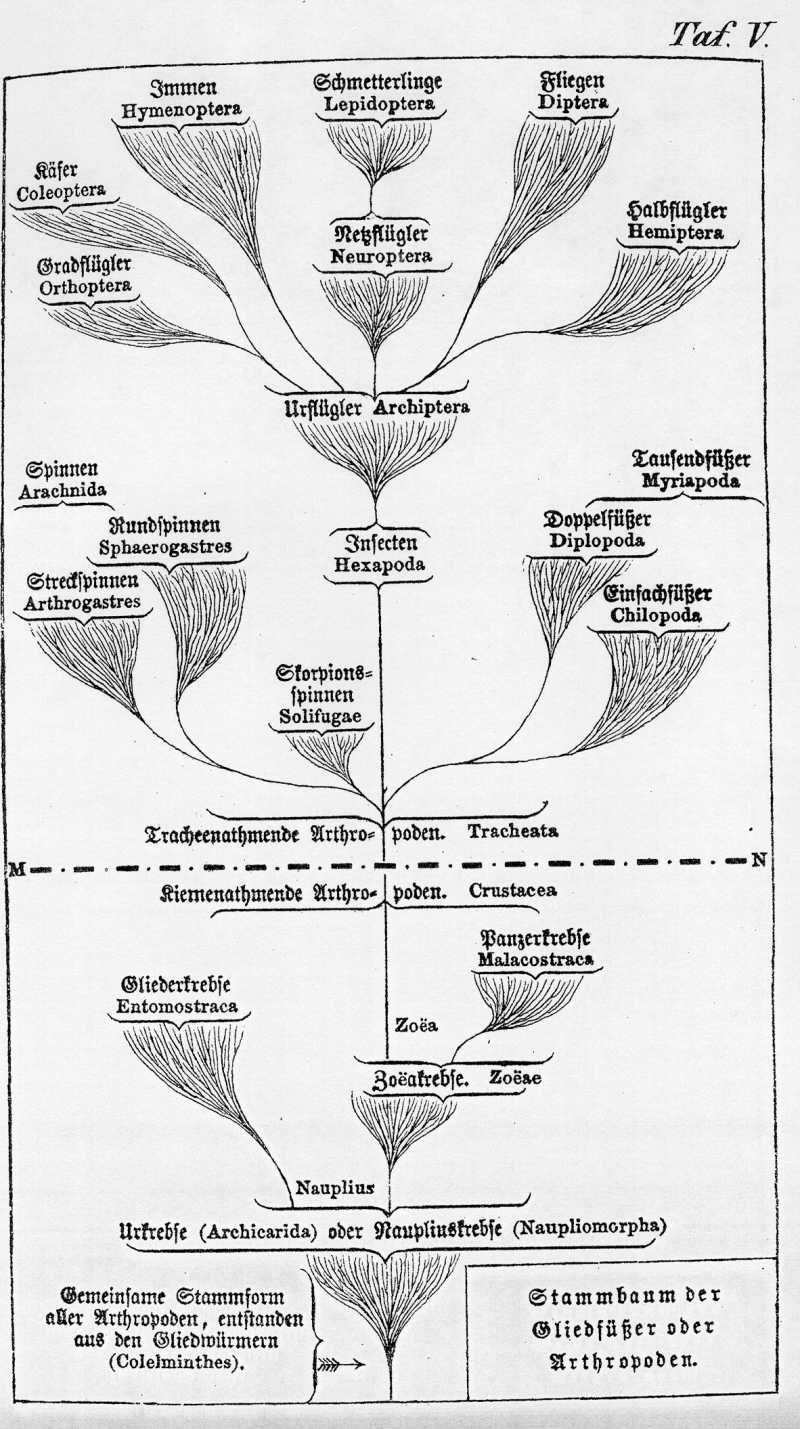
Von den Sternthieren wenden wir uns zu dem fünften und höchst entwickelten Stamm unter den wirbellosen Thieren, zu dem Phylum der Gliedfüßer (Arthropoda). Wie schon vorher bemerkt wurde, entspricht dieser Stamm der Klasse der Kerfe oder Insecten im ursprünglichen Sinne Linné's. Er enthält wiederum vier Klassen, nämlich 1. die echten sechsbeinigen Insecten; 2. die achtbeinigen Spinnen; 3. die mit zahlreichen Beinpaaren versehenen Tausendfüße und 4. die mit einer wechselnden Beinzahl versehenen Krebse oder Krustenthiere. Die letzte Klasse athmet Wasser durch Kiemen und kann daher als Hauptklasse der kiemenathmenden Arthropoden oder Kiemenkerfe (Carides) den drei ersten Klassen entgegengesetzt werden. Diese athmen Luft durch durch eigenthümliche Luftröhren oder Tracheen, und können daher passend in der Hauptklasse der tracheenathmenden Arthropoden oder Tracheenkerfe (Tracheata) vereinigt werden.
Bei allen Gliedfüßern sind, wie der Name sagt, die Beine deutlich gegliedert, und dadurch, sowie durch die stärkere Differenzirung der gegliederten Körperabschnitte oder Metameren unterscheiden sie sich wesentlich von den Würmern, mit denen sie Bär und Cuvier in dem Typus der Gliederthiere oder Articulaten vereinigten. Uebrigens stehen sie den Gliedwürmern (Colelminthes) in jeder Beziehung so nahe, daß sie kaum scharf von ihnen zu trennen sind. Insbesondere theilen sie mit den Ringelwürmern die sehr charakteristische Form des centralen Nervensystems, das sogenannte Bauchmark, welches vorn mit einem den Mund umgebenden Schlundring beginnt. Auch aus anderen Thatsachen geht hervor, daß die Arthropoden sich jedenfalls aus Gliedwürmern erst später entwickelt haben. Wahrscheinlich sind die Räderthiere und demnächst die Ringelwürmer ihre nächsten Blutsverwandten im Würmerstamme (Gen. Morph. II., Taf. V., S. LXXXV-CII).
Der Stammbaum der Arthropoden läßt sich aus der Paläontologie, vergleichenden Anatomie und Ontogenie seiner vier Klassen in seinen Grundzügen vortrefflich erkennen, obwohl auch hier, wie überall, im Einzelnen noch sehr Vieles dunkel bleibt. Die Wurzel des ganzen Phylum bildet die Klasse der Kiemenkerfe oder Krebse (Carides), wegen ihrer harten krustenartigen Körperbedeckung auch Krustenthiere (Crustacea) genannt. Die Ontogenie oder die individuelle Entwickelungsgeschichte der Krebse ist außerordentlich interessant, und verrräth uns, ebenso wie bei den Wirbelthieren, deutlich die wesentlichen Grundzüge ihrer Stammesgeschichte oder Phylogenie. Fritz Müller hat in seiner ausgezeichneten, bereits angeführten Schrift "Für Darwin"16) dieses merkwürdige Verhältniß vortrefflich erläutert.
Die gemeinschaftliche Stammform aller Krebse, welche sich bei den meisten noch heutzutage zunächst aus dem Ei entwickelt, ist ursprünglich ein und dieselbe: der sogenannte Nauplius. Dieser merkwürdige Urkrebs ist eine sehr einfache gegliederte Thierform, welche sich zunächst an die Räderthiere anschließt und aus ähnlichen Gliedwürmern wahrscheinlich seinen Ursprung genommen hat. Aus der gemeinsamen Larvenform des Nauplius entwickeln sich als divergente Zweige nach verschiedenen Richtungen hin die sechs Ordnungen der niederen Krebse, welche in der nachstehenden systematischen Uebersicht des Arthropodenstammes als Gliederkrebse (Entomostraca) zusammengefaßt sind. Auch die höhere Abtheilung der Panzerkrebse (Malacostraca) hat aus der gemeinsamen Naupliusform ihren Ursprung genommen. Jedoch hat sich hier der Nauplius zunächst in eine andere Larvenform, die sogenannte Zo'a, umgewandelt, welche eine außerordentliche Bedeutung besitzt. Diese seltsame {Tabelle Seite 424} Zo'a ist nämlich aller Wahrscheinlichkeit nach nicht allein die gemeinsame Stammform für alle sechs beistehend verzeichneten Ordnungen der Malacostraca, sondern auch zugleich für die luftathmenden Tracheenkerfe, für die Spinnen, Tausendfüße und Insecten.

Diese letzteren sind jedenfalls erst im Angang der paläolitischen Zeit, nach Abschluß des archolithischen Zeitraums entstanden, weil alle diese Thiere (im Gegensatz zu den meist wasserbewohnenden Krebsen) ursprünglich Landbewohner sind. Offenbar können sich diese Luftathmer erst entwickelt haben, als nach Verfluß der silurischen Zeit das Landleben begann. Da nun aber fossile Reste von Spinnen und Insekten bereits in den Steinkohlenschichten gefunden werden, so können wir ziemlich genau den Zeitpunkt ihrer Entstehung feststellen. Es muß die Entwickelung der ersten Tracheenkerfe aus kiemenathmenden Zo'akrebsen zwischen das Ende der Silurzeit und den Beginn der Steinkohlenzeit fallen, also entweder in die antedevonische oder in die devonische oder in die antecarbonische Periode.
Die gemeinschaftliche Ausgangsform der drei durch Tracheen athmenden Arthropodenklassen ist uns wahrscheinlich bis auf den heutigen Tag nur wenig verändert in einer merkwürdigen Spinnenform erhalten. Diese uralte Tracheatenform ist die Skorpionsspinne (Solifuga), von der mehrere große, wegen ihres giftigen Bisses sehr gefürchtete Arten noch heute im wärmeren Asien leben. Der Körper besteht hier, wie wir es bei dem gemeinsamen Stammvater der Tracheaten voraussetzen müssen, aus drei getrennten Abschitten, einem Kopfe, welcher mehrere beinartige Kieferpaare trägt, einer Brust, an deren drei Ringen drei Beinpaare befestigt sind, und einem anhangslosen Hinterleib. Wahrscheinlich haben sich aus unbekannten devonischen Tracheaten, als zwei divergente Aeste einerseits die echten Spinnen, andrerseits die Insecten entwickelt. Die Tausendfüßer sind entweder ein eigenthümlich entwickelter Seitenzweig der Insecten oder ein dritter Ast jener Stammform.
Die echten Spinnen (Arachnida) sind durch den Mangel der Flügel und durch vier Beinpaare von den Insecten unterschieden. Wie jedoch die Skorpionspinnen und die Taranteln deutlich zeigen, sind eigentlich auch bei ihnen, wie bei den Insecten, nur drei echte Beinpaare vorhanden. Das scheinbare vierte Beimpaar der Spinnen (das vorderste) ist eigentlich ein Kieferfußpaar.
Die Spinnenklasse zerfällt in zwei Unterklassen: Streckspinnen und Rundspinnen.
Von diesen sind die Streckspinnen (Arthrogastres) die älteren und ursprünglichen Formen, bei denen sich die frühere Leibesgliederung besser erhalten hat.
Es gehören dahin außer den schon genannten Skorpionsspinnen (Solifugae) und den Taranteln (Phrynida) die gefürchteten echten Skorpione (Scorpioda), die kleinen, in unseren Bibliotheken wohnenden Bücherskorpione (Pseudoscorpioda), die langbeinigen Schneiderspinnen (Opiliones) und die im Meere lebenden seltsamen Asselspinnen (Pycnogonida).
Versteinerte Reste von Streckspinnen finden sich bereits in der Steinkohle. Dagegen kommt die zweite Unterklasse der Arachniden, die Rundspinnen (Sphaerogastres) versteinert zuerst im Jura, also sehr viel später vor. Sie haben sich aus einem Zweige der Streckspinnen dadurch entwickelt, daß die Leibesringe mehr oder weniger mit einander verschmolzen. Bei den eigentlichen Webespinnen (Araneae), welche wir wegen ihrer feinen Webekünste bewundern, geht die Verschmelzung der Rumpfglieder oder Metameren so weit, daß der Rumpf nur noch aus zwei Stücken besteht, einer Kopfbrust, welche die Kiefer und die vier Beinpaare trägt, und einem anhangslosen Hinterleib, an welchem die Spinnwarzen sitzen.
Bei den Milben (Acara), welche wahrscheinlich aus einem verkümmerten Seitenzweige der Webespinnen durch Entartung (insbesondere durch Schmarotzerleben) entstanden sind, verschmelzen sogar noch diese beiden Rumpfstücke mit einander zu einer ungegliederten Masse.
Die Klasse der Tausendfüßer (Myriapoda)., die kleinste und formenärmste unter den vier Arthropodenklassen, zeichnet sich durch den sehr verlängerten Leib aus, welcher einem gegliederten Ringelwurme sehr ähnlich ist und oft mehrere hundert Beinpare trägt. Aber auch sie hat sich ursprünglich aus einer sechsbeinigen Tracheatenform entwickelt, wie die individuelle Entwickelung der Tausendfüßer im Eie deutlich beweist. Ihr Embryonen haben zuerst nur drei Beinpaare, gleich den echten Insecten, und erst später knospen Stück für Stück die folgenden Beinpaare aus den wuchernden Hinterleibsringen hervor. Von den beiden Ordnungen der Tausendfüßer (welche bei uns unter Baumrinden, im Mose u. s. w. leben), haben sich wahrscheinlich die runden Doppelfüßer (Diplopoda) erst später aus den alten platten Einfachfüßern (Chilopoda) entwickelt. Von den letzteren finden sich fossile Reste zuerst im Jura vor.
Die dritte und letzte Klasse unter den tracheenathmenden Arthropoden ist die der Insecten (Insecta oder Hexapoda), die umfangreichste von allen Thierklassen, und nächst derjenigen der Säugethiere auch die wichtigste von allen. Trotzdem die Insecten eine größere Mannichfaltigkeit von Gattungen und Arten entwickelten, als die übrigen Thiere zusammengenommen, sind das alles doch im Grunde nur oberflächliche Variationen eines einzigen Themas, welches in seinen wesentlichen Charakteren sich ganz beständig erhält. Bei allen Insecten sind die drei Abschnitte des Rumpfes, Kopf, Brust und Hinterleib deutlich getrennt. Der Hinterleib oder das Abdomen trägt, wie bei den Spinnen, gar keine gegliederten Anhänge.
Der mittlere Abschnitt, die Brust oder der Thorax trägt auf der Bauchseite die drei Beinpaare, auf der Rückenseite ursprünglich zwei Flügelpaare. Freilich sind bei sehr vielen Insecten eines oder beide Flügelpaare verkümmert, oder selbst ganz verschwunden. Allein die vergleichende Anatomie der Insecten zeigt uns deutlich, daß dieser Mangel erst nachträglich durch Verkümmerung der Flügel entstanden ist, und daß alle (oder doch die meisten) jetzt lebenden Insecten von einem gemeinsamen Stamminsect abstammen, welches drei Beinpaare und zwei Flügelpaare besaß (Vergl. S. 233). Diese Flügel, welche die Insecten so auffallend vor den übrigen Gliedfüßern auszeichnen, entstanden wahrscheinlich aus den Tracheenkiemen, welche wir noch heute an den im Wasser lebenden Larven der Eintagsfliegen (Ephemera) beobachten.
Der Kopf der Insecten trägt allgemein außer den Augen ein Paar gegliederte Fühlhörner oder Antennen, und außerdem auf jeder Seite des Mundes drei Kiefer. Diese drei Kieferpaare, obgleich bei allen Insecten aus derselben ursprünglichen Grundlage entstanden, haben sich durch verschiedenartige Anpassung bei den verschiedenen Orndungen zu höchst mannichfaltigen und merkwürdigen Formen umgebildet, so daß man sie hauptsächlich zur Unterscheidung und Charakteristik der Hauptabtheilungen der Klasse verwendet.
Zunächst kann man als zwei Hauptabtheilungen Insecten mit kauenden Mundtheilen (Masticantia) und Insecten mit saugenden Mundtheilen (Sugentia) unterscheiden. Bei genauerer Betrachtung kann man noch schärfer jede dieser beiden Abtheilungen in zwei Untergruppen vertheilen. Unter den Kauinsecten oder Masticantien können wir die beißenden und die leckenden unterscheiden. Zu den Beißenden (Mordentia) gehören die ältesten und ursprünglichsten Insecten, die vier Ordnungen der Urflügler, Netzflügler, Gradflügler und Käfer. Die Leckenden (Lambentia) werden bloß durch die eine Ordnung der Hautflügler gebildet. Unter den Sauginsecten oder Sugentien können die beiden Gruppen der stechenden und schlürfenden unterscheiden. Zu den Stechenden (Pungentia) gehören die beiden Ordnungen der Halbflügler und Fliegen, zu den Schlürfenden (Sorbentia) bloß die Schmetterlinge.
Als die ältesten Insecten, welche sich aus unbekannten, den Skorpionsspinnen ähnlichen Arachniden entwickelten, betrachte ich die beißenden, und zwar die Ordnung der Urflügler (Archiptera oder Pseudoneuroptera). Dahin gehöhren vor allen die Eintagsfliegen (Ephemera), deren im Wasser lebende Larven uns wahrscheinlich noch heute in ihren Tracheenkiemen die Organe zeigen, aus denen die Insectenflügel ursrünglich entstanden. Ferner gehören in diese Ordnung die bekannten Wasserjungfern oder Libellen, die flügellosen Zuckergäste (Lepisma), die springenden Blasenfüßer (Physopoda), und die gefürchteten Termiten, von denen sich versteinerte Reste schon in der Steinkohle finden. Unmittelbar hat sich wahrscheinlich aus den Urflüglern die Ordnung der Netzflügler (Neuroptera) entwickelt, welche sich von ihnen wesentlich nur durch die vollkommene Verwandlung unterscheiden. Es gehören dahin die Florfliegen (Planipennia), die Schmetterlingsfliegen (Phryganida) und die Fächerfliegen (Strepsiptera). Fossile Insecten, welche den Uebergang von den Urflüglern (Libellen) zu den Netzflüglern (Sialiden) machen, kommen schon in der Steinkohle vor (Dictyophlebia). Aus einem anderen Zweige der Urflügler hat sich durch Differenzirung der beiden Flügelpaare schon frühzeitig die Ordnung der Gradflügler (Orthoptera) entwickelt. Diese Abtheilung besteht aus der formenreichen Gruppe der Schaben, Heuschrecken, Gryllen u. s. w. (Ulonata, und aus der kleinen Gruppe der bekannten Ohrwürmer (Labidura), welche durch die Kneifzange am hinteren Körperende ausgezeichnet sind. Sowohl von Schaben als von Gryllen und Heuschrecken kennt man Versteinerungen aus der Steinkohle.
Auch die vierte Ordnung der beißenden Insecten, die Käfer (Coleoptera) kommen bereits in der Steinkohle versteinert vor. Diese außerordentlich umfangreiche Ordnung, der bevorzugte Liebling der Insectenliebhaber und Sammler, zeigt am deutlichsten von allen, welche unendliche Formenmannichfaltigkeit sich durch Anpassung an verschiedene Lebensverhältnisse äußerich entwickeln kann, ohne daß deshalb der innere Bau und die Grundform des Körpers irgendwie wesentlich umgebilet wird. Wahrscheinlich haben sich die Käfer aus einem Zweige der Gradflügler entwickelt, von denen sie sich wesenlich nur durch ihre vollkommene Verwandlung unterscheiden.
An diese vier Ordnungen der beißenden Insecten schließt sich nun zunächst die eine Ordnung der leckenden Insecten an, die interessante Gruppe der Immen oder Hautflügler (Hymenoptera). Dahin gehören diejenigen Insecten, welche sich durch ihre entwickelten Culturzustände, durch ihre weitgehende Arbeitstheilung, Gemeindebildung und Staatenbildung zu bewunderungswürdiger Höhe der Geistesbildung, der intellectuellen Vervollkommnung und der Charakterstärke erhoben haben und dadurch nicht allein die meisten Wirbellosen, sondern überhaupt die meisten Thiere übertreffen. Es sind das vor allen die Ameisen und die Bienen, sodann die Wespen, Blattwespen, Holzwespen, Schlupfwespen, Gallwespen u. s. w. Sie kommen versteinert im Jura vor, in größerer Menge jedoch erst in der Tertiärschichten. Wahrscheinlich haben sich die Hautflügler aus einem Zweige entweder der Urflügler oder der Netzflügler entwickelt.
Von den beiden Ordnungen der stechenden Insecten, den Hemipteren und Dipteren, ist die ältere diejenige der Halbflügler (Hemiptera), auch Schnabelkerfe (Rhynchota) genannt. Dahin gehören die drei Unterordnungen der Blattläuse (Homoptera), der Wanzen (Heteroptera) und der Läuse (Pediculina). Von ersteren beiden finden sich fossile Reste schon im Jura. Aber schon im permischen System kommt ein altes Insect vor (Eugereon), welches auf die Abstmmung der Hemipteren von den Neuropteren hinzudeuten scheint. Wahrscheinlich sind von den drei Unterordnungen der Hemipteren die ältesten die Homopteren, zu denen außer den eigentlichen Blattläusen auch noch die Schildläuse, die Blattflöhe und die Zirpen oder Cicaden gehören. Aus zwei verschiedenen Zweigen der Homopteren werden sich die Läuse durch weitgehende Entartung (vorzüglich Verlust der Flügel), die Wanzen dagegen durch Vervollkommnung (Sonderung der beiden Flügelpaare) entwickelt haben.
Die zweite Ordnung der stechenden Insecten, die Fliegen oder Zweiflügler (Diptera) findet sich zwar auch schon im Jura versteinert neben den Halbflüglern vor. Allein dieselben haben sich doch wahrscheinlich erst nachträglich aus den Hemipteren durch Rückbildung der Hinterflügel entwickelt. Nur die Vorderflügel sind bei den Dipteren vollständig entwickelt. Die Hauptmasse dieser Ordnung bilden die langgesteckten Mücken (Nemocera) und die gedrungenen eigentlichen Fliegen (Brachycera), von denen die ersteren wohl älter sind. Doch finden sich von Beiden schon Reste im Jura vor. Durch Degeneration in Folge von Parasitismus haben sich aus ihnen wahrscheinlich die beiden kleinen Gruppen der puppengebärenden Lausfliegen (Pupipara) und der springenden Flöhe (Aphaniptera) entwickelt.
Die achte und letzte Insectenordnung, und zugleich die einzige mit wirklich schlürfenden Mundtheilen sind die Schmetterlinge (Lepidoptera). Diese Ordnung erscheint in mehreren morphologischen Beziehungen als die vollkommenste Abtheilung der Insecten und hat sich demgemäß auch am spätesten erst entwickelt. Man kennt nämlich von dieser Ordnung Versteinerungen nur aus der Tertiärzeit, während die drei vorhergehenden Ordnungen bis zu Jura, die vier beißenden Ordnungen dagegen sogar bis zur Steinkohle hinaufreichen. Die nahe Verwandtschaft einiger Motten (Tinea) und Eulen (Noctua) mit einigen Schmetterlingsfliegen (Phryganida) macht es wahrscheinlich, daß sich die Schmetterlinge aus dieser Gruppe, also aus der Ordnung der Netzflügler oder Neuroptheren entwickelt haben.
Wie Sie sehen, bestätigt Ihnen die ganze Geschichte der Insectenklasse und weiterhin auch die Geschichte des ganzen Arthropodenstammes wesentlich die großen Gesetze der Differenzirung und Vervollkommnung, welche wir nach Darwin's Selectionstheorie als die nothwendigen Folgen der natürlichen Züchtung anerkennen müssen. Der ganze formenreiche Stamm beginnt in archolithischer Zeit mit der kiemenathmenden Klasse der Krebse, und zwar mit den niedersten Urkrebsen oder Archicariden. Die Gestalt dieser Urkrebse, die sich jedenfalls aus Gliedwürmern, und zwar wahrscheinlich aus Räderthieren entwickeltn, ist uns noch heute in der gemeinsamen Jugendform aller niederen oder Gliederkrebse (Entomostraca), in dem merkwürdigen Nauplius, annähernd erhalten. Aus dem Nauplius entwickelte sich weiterhin die seltsäme Zo'a, die gemeinsame Jugendform aller höheren oder Panzerkrebse (Malacostraca) und zugleich desjenigen, zuerst durch Tracheen athmenden Arthropoden, welcher der gemeinsame Stammvater aller Tracheaten wurde. Dieser Stammvater, der zwischen dem Ende der Silurzeit und dem Beginn der Steinkohlenzeit entstanden sein muß, stand wahrscheinlich von allen jetzt noch lebenden Insecten den Skorpionsspinnen oder Solifugen am nächsten. Aus ihm entwickelten sich aus drei divergenten Zweige die drei Tracheaten Klassen, Spinnen, Tausendfüßer und echte (sechsbeiige und vierflügelige) Insecten. Von diesen letzteren existirten lange Zeit hindurch nur die vier beißenden Ordnungen, Urflügler, Netzflügler, Gradflügler und Käfer, von denen die erste wahrscheinlich die gemeinsame Stammform der drei anderen ist. Erst viel später entwickelten sich aus den beißenden Insecten, welche die ursprüngliche Form der drei Kieferpaare am reinsten bewahrten, als drei divergente Zweige die leckenden, stechenden und schlürfenden Insecten. Wie diese Ordnungen in der Erdgeschichte auf einander folgten, zeigt Ihnen nochmals über sichtlich die nachstehende Tabelle. {Tabelle Seite 432 }

Anmerkung: Bei den acht einzelnen Ordnungen der Insecten ist zugleich der Unterschied in der Metamorphose oder Verwandlung und in der Flügelbildung durch folgende Buchstaben angegeben: M. I. = Unvollständige Metamorphose. M. C. = Vollständige Metamorphose (Vergl. Gen. Morph. II, S. XCIX). A. A. = Gleichartige Flügel (Vorder- und Hinterflügel im Bau und Gewebe nicht oder nur wenig verschieden). A. D. = Ungleichartige Flügel (Vorder- und Hinterflügel durch starke Differenzirung im Bau und Gewebe sehr verschieden).
Stammbaum und Geschichte des Pflanzenreichs

 Stammbaum und Geschichte des Thierreichs. II.
Stammbaum und Geschichte des Thierreichs. II.
Renaissance Forum - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - erstes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Jacob Burckhardt - Die Kultur der Renaissance in Italien. Ein Versuch
Leonardo da Vinci Wissenschaftler - Erfinder - Künstler
Venedig - Eine Liebeserklärung an eine Stadt
William Shakespeare animiert (wahrscheinlich oder zufällig...2/3 zu 1/3) William Turner
Philosophie für Schnelldenker - Besinnliche Philosophie
Startseite - © Copyright 2004- - Susanne Albers - Kiehlufer 125 - D 12059 Berlin