Startseite - Impressum - Datenschutzerklärung - News - Ich diene dem, der hört - https://offenunddirekt.de - Mein Gratis-Coaching für Dich - Kraft tanken - Spirituelles - Kunst - Technik - Literatur - Kultur - Politik - Architektur - Spaß - Psychologie - Spiele - Fotos + Animationen - Gästebuch - Persönliches - Blog - Die Stimme Gottes - mein Youtube Kanal - erstes Rätsel - Renaissance Forum - Chat - Quiz - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - Zufallsspiel - letztes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Ernst Haeckel - Die Natürliche Schöpfungsgeschichte 1868
Fünfzehnter Vortrag.
Stammbaum und Geschichte des Protistenreichs.
(Hierzu Taf. I.)
Meine Herren! Durch die denkende Vergleichung der individuellen und paläontologischen Entwickelung, sowie durch die vergleichende Anatomie der Organismen, durch die vergleichende Betrachtung ihrer entwickelten Formverhältnisse, gelangen wir zur Erkenntniß ihrer stufenweis verschiedenen Formverwandtschaft. Dadurch gewinnen wir aber zugleich einen Einblick in ihre wahre Blutsverwandtschaft, welche nach der Descendenztheorie der eigentliche Grund der Formverwandtschaft ist. Wir gelangen also, indem wir die empirischen Resultate der Embryologie, Paläontologie und Anatomie zusammenstellen, vergleichen, und zur gegenseitigen Ergänzung benutzen, zur annähernden Erkenntniß des natürlichen Systems, welches nach unserer Ansicht der Stammbaum der Organismen ist. Allerdings bleibt unser menschliches Wissen, wie überall, so ganz besonders hier, nur Stückwerk, schon wegen der außerordentlichen Unvollständigkeit und Lückenhaftigkeit der empirischen Schöpfungsurkunden. Indessen dürfen wir uns dadurch nicht abschrecken lassen, jene höchste Aufgabe der Biologie in Angriff zu nehmen. Lassen Sie uns vielmehr sehen, wie weit es schon jetzt möglich ist, trotz des unvollkommenen Zustandes unserer embryologischen, paläontologischen und anatomischen Kenntnisse, eine annähernde Hypothese von dem verwandtschaftlichen Zusammenhang der Organismen aufzustellen.
Darwin gibt uns in seinem Werk auf diese speciellen Fragen der Descendenztheorie keine Antwort. Er äußert nur am Schlusse derselben seine Vermuthung, "daß die Thiere von höchstens vier oder fünf, und die Pflanzen von eben so vielen oder noch weniger Stammarten herrühren." Da aber auch diese wenigen Hauptformen noch Spuren von verwandtschaftlicher Verkettung zeigen, und da selbst Pflanzen- und Thierreich durch vermittelnde Uebergangsformen verbunden sind, so gelangt er weiterhin zu der Annahme, "daß wahrscheinlich alle organischen Wesen, die jemals auf dieser Erde gelebt, von irgend einer Urform abstammen, welcher das Leben zuerst vom Schöpfer eingehaucht worden ist." Gleich Darwin haben auch alle anderen Anhänger der Descendenztheorie dieselbe bloß im Allgmeinen gefördert, und nicht den Versuch gemacht, sie auch speciell durchzuführen, und das "natürliche System" wirklich als "Stammbaum der Organismen" zu behandeln. Wenn wir daher hier dieses schwierige Unternehmen wagen, so müssen wir uns ganz auf unsere eigene Füße stellen.
Ich habe vor zwei Jahren in der systematischen Einleitung zu meiner allgemeinen Entwickelungsgeschichte (im zweiten Bande der generellen Morphologie) eine Anzahl von hypothetischen Stammtafeln für die größeren Organismengruppen aufgestellt, und damit thatsächlich den ersten Versuch gemacht, die Stammbäume der Organismen in der Weise, wie es die Entwickelungstheorie erfordert, wirklich zu construiren. Dabei war ich mir der außerordentlichen Schwierigkeiten dieser Aufgabe vollkommen bewußt. Indem ich trotz aller abschreckenden Hindernisse dieselbe in Angriff nahm, beanspruchte ich weiter Nichts als den ersten Versuch gemacht und zu weiteren und besseren Versuchen angeregt zu haben. Vermuthlich werden die meisten Zoologen und Botaniker von diesem Anfang sehr wenig befriedigt gewesen sein, und am wenigsten in dem engen Specialgebiete, in welchem ein Jeder besonders arbeitet. Allein wenn irgendwo, so ist ganz gewiß hier das Tadeln viel leichter als das Bessermachen, und daß bisher noch kein Naturforscher meine Stammbäume durch bessre oder überhaupt durch andere ersetzt hat, beweist am besten die ungeheure Schwierigkeit der unendlich verwickelten Aufgabe. Aber gleich allen anderen wissenschaftlichen Hypothesen, welche zur Erklärung der Thatsachen dienen, werden auch meine genealogischen Hypothesen so lange auf Berücksichtigung Anspruch haben, bis sie durch bessere ersetzt werden.
Hoffentlich wird dieser Ersatz recht bald geschehen, und ich wünschte Nichts mehr, als daß mein erster Versuch recht viele Naturforscher anregen möchte, wenigstens auf dem engen, ihnen genau bekannten Specialgebiete des Thier- oder Pflanzenreichs die genaueren Stammbäume für einzelne Gruppen aufzustellen. Durch zahlreiche derartige Versuche wird unsere genealogische Erkenntniß im Laufe der Zeit langsam fortschreiten, und mehr und mehr der Vollendung näher kommen, obwohl mit Bestimmtheit vorauszusehen ist, daß ein vollendeter Stammbaum niemals wird erreicht werden. Es fehlen uns und werden uns immer fehlen die unerläßlichen paläontologischen Grundlagen. Die ältesten Urkunden werden uns ewig verschlossen bleiben aus den früher bereits angeführten Ursachen. Die ältesten, durch Urzeugung entstandenen Organismen, die Stammeltern aller folgenden, müssen wir uns nothwendig als Moneren denken, als einfache weiche Eiweißklümpchen, ohne jede bestimmte Form, ohne irgend welche harte Theile. Diese waren daher der Erhaltung im versteinerten Zustande, durchaus nicht fähig.
Ebenso fehlt uns aber aus den im letzten Vortrage ausführlich erörterten Gründen der bei weiten größte Theil von den zahllosen paläontologischen Dokumenten, die zur Durchführung der Stammesgeschichte oder Phylogenie, und zur wahren Erkenntniß der organischen Stammbäume eigentlich erforderlich wären. Wenn wir daher das Wagniß ihrer hypothetischen Construction dennoch unternehmen, so sind wir vor Allem auf die Unterstützung der beiden anderen Urkundenreihen hingewiesen, welche das paläontologische Archiv in wesentlicher Weise ergänzen, der Ontogenie und der vergleichenden Anatomie.
Ziehen wir diese höchst werthvollen Urkunden gehörig denkend und vergleichend zu Rathe, so machen wir zunächst die außerordentlich bedeutungsvolle Wahrnehmung, daß die allermeisten Organismen, insbesondere alle höheren Thiere und Pflanzen, aus einer Vielzahl von Zellen zusammengesetzt sind, ihren Ursprung aber aus einem Ei nehmen, und daß dieses Ei bei den Theren ebenso wie bei den Pflanzen eine einzige ganz einfache Zelle ist: ein Klümpchen einer Eiweißverbindung, in welchem ein anderer eiweißartiger Körper, der Zellkern, eingeschlossen ist.
Diese kernhaltige Zelle wächst und vergrößert sich. Durch Theilung bildet sie ein Zellenhäufchen, und aus diesem entstehen durch Arbeitstheilung in der früher beschriebenen Weise die vielfach verschiedenen Formen, welche die ausgebildeten Thier- und Pflanzenarten uns vor Augen führen. Dieser unendlich wichtige Vorgang, welchen wir alltäglich bei der embryologischen Entwickelung jedes thierischen und pflanzlichen Individuums mit unseren Augen Schritt für Schritt unmittelbar verfolgen können, und welchen wir in der Regel durchaus nicht mit der verdienten Ehrfurcht betrachten, belehrt uns sicherer und vollständiger, als alle Versteinerungen es thun könnten, über die ursprüngliche paläontologische Entwickelung aller mehrzelligen Organismen, aller höheren Thiere und Pflanzen.
Denn da die Ontogenie oder die embryologische Entwickelung jedes einzelnen Individuums Nichts weiter ist als eine Recapitulation der Phylogenie oder der paläontologischen Entwickelung seiner Vorfahrenkette, so können wir daraus zunächst mit voller Sicherheit den ebenso einfachen als bedeutenden Schluß ziehen, daß alle mehrzelligen Thiere und Pflanzen ursprünglich von einzelligen Organismen abstammen. Die uralten primordialen Vorfahren des Menschen so gut wie aller anderen Thiere und aller aus vielen Zellen zusammengesetzten Pflanzen waren einfache, isolirt lebende Zellen. Dieses unschätzbare Geheimniß des organischen Stammbaums wird uns durch das Ei der Thiere und durch das "Keimbläschen" der Pflanzen mit untrüglicher Sicherheit verrathen. Wenn die Gegner der Discendenztheorie uns entgegenhalten, es sei wunderbar und unbegreiflich, daß ein äußerst complicirter vielzelliger Organismus aus einem einzelligen Organismus im Laufe der Zeit hervorgegangen sei, so entgegnen wir einfach, daß wir dieses unglaubliche Wunder jeden Augenblick vor uns sehen und mit unseren Augen verfolgen können. Denn die Embryologie der Thiere und Pflanzen führt uns in kürzester Zeit denselben Vorgang greifbar vor Augen, welcher im Laufe ungeheurer Zeiträume bei der Entstehung des ganzen Stammes stattgefunden hat.
Auf Grund der embryologischen Urkunden können wir also mit voller Sicherheit behaupten, daß alle mehrzelligen Organismen eben so gut wie alle einzelligen ursprünglich von einfachen Zellen abstammen; hieran würde sich sehr natürlich der Schluß reihen, daß die älteste Wurzel des Thier- und Pflanzenreichs gemeinsam ist. Denn die verschiedenen uralten "Stammzellen", aus denen sich die wenigen verschiedenen Hauptgruppen oder "Stämme" (Phylen) des Thier- und Pflanzenreichs entwickelt haben, könnten ihre Verschiedenheit selbst erst erworben haben, und könnten selbst von einer gemeinsamen "Urstammzelle" abstammen. Wo kommen aber jene wenigen "Stammzellen" oder diese "Urstammzelle" her? Zur Beantwortung dieser genealogischen Grundfrage müssen wir auf die früher erörterte Plastidentheorie und die Urzeugungstheorie zurückgreifen.
Wie wir damals zeigten, können wir uns durch Urzeugung unmittelbar nicht Zellen entstanden denken, sondern nur Moneren, Urwesen der denkbar einfachsten Art, gleich den noch jetzt lebenden Protamoeben, Protomyxen, Protogenes u. s. w. (S. 144, Fig. 1). Nur solche structurlose Schleimkörperchen, deren ganzer eiweißartiger Leib so homogen wie ein anorganischer Krystall ist, und die dennoch die beiden organischen Grundfunctionen der Ernährung und Fortpflanzung vollziehen, konnten unmittelbar im Beginn der antelaurentischen Zeit aus anorganischer Materie durch Autogenie entstehen. Während einige Moneren auf der ursprünglichen einfachen Bildungsstufe verharrten, bildeten sich andere allmählich zu Zellen um, indem der innere Kern des Eiweißleibes sich von dem äußeren Zellstoff sonderte. Andrerseits bildete sich durch Differenzirung der äußersten Zellstoffschicht sowohl um einfache (kernlose) Cytoden, als um nackte (aber kernhaltige) Zellen eine äußere Hülle (Membran oder Schale). Durch diese beiden Sonderungsvorgänge in dem einfachen Urschleim des Monerenleibes, durch die Bildung eines Kerns im Inneren, einer Hülle an der äußeren Oberfläche des Plasmakörpers, entstanden aus den ursprünglichen einfachsten Cytoden, den Moneren, jene vier verschiedenen Arten von Plastiden oder Individuen erster Ordnung, aus denen weiterhin alle übrigen Organismen durch Differenzirung und Zusammensetzung sich entwickeln konnten. (Vergl. oben S. 286).
Hier wird sich Ihnen nun zunächst die Frage aufdrängen: Stammen alle organischen Cytoden und Zellen, und mithin auch jene Stammzellen, welche wir vorher als die Stammeltern der wenigen großen Hauptgruppen des Thier- und Pflanzenreichs betrachtet haben, von einer einzigen ursprünglichen Monerenform ab, oder giebt es mehrere verschiedene organische Stämme, deren jeder von einer eigenthümlichen, selbstständig durch Urzeugung entstandenen Monerenart abzuleiten ist. Mit anderen Worten: Ist die ganze organische Welt gemeinsamen Ursprungs, oder verdankt sie mehrfachen Urzeugungsakten ihre Entstehung? Diese genealogische Grundfrage scheint auf den ersten Blick ein außerordentliches Gewicht zu haben. Indessen werden Sie bei näherer Betrachtung bald sehen, daß sie dasselbe nicht besitzt, vielmehr im Grunde von sehr untergeordneter Bedeutung ist.
Lassen Sie uns hier zunächst den Begriff des organischen Stammes näher in's Auge fassen und fest begrenzen. Wir verstehen unter Stamm oder Phylum die Gesammtheit aller derjenigen Organismen, deren Blutsverwandtschaft, deren Abstammung von einer gemeinsamen Stammform aus anatomischen und entwicklungsgeschichtlichen Gründen nicht zweifelhaft sein kann, oder doch wenigstens in hohem Maße wahrscheinlich ist. Unsere Stämme oder Phylen fallen also wesentlich dem Begriffe nach zusammen mit jenen wenigen "großen Klassen" oder "Hauptklassen," von denen Darwin glaubt, daß eine jede nur blutsverwandte Organismen enthält, und von denen er sowohl im Thierreich als im Pflanzenreich nur sehr wenige, in jedem Reiche etwa vier bis fünf annimmt. Im Thierreich würden diese Stämme im Wesentlichen mit jenen vier bis sechs Hauptabtheilungen zusammenfallen, welche die Zoologen seit Bär und Cuvier als "Hauptformen, Generalpläne, Zweige oder Kreise" des Thierreichs unterscheiden (Vgl. S. 42). Bär und Cuvier unterschieden deren nur vier, nämlich 1. die Wirbelthiere (Vertebrata); 2. die Gliederthiere (Articulata); 3. die Weichthiere (Mollusca) und 4. die Strahlthiere (Radiata).
Gegenwärtig unterscheidet man gewöhnlich sechs, indem man den Stamm der Gliederthiere in die beiden Stämme der Gliederfüßer (Arthropoda) und der Würmer (Vermes) trennt, und ebenso den Stamm der Strahlthiere in die beiden Stämme der Sternthiere (Echinoderma) und der Pflanzenthiere (Coelenterata) zerlegt. Innerhalb jedes dieser sechs Stämme zeigen alle dazu gehörigen Thiere trotz großer Mannichfaltigkeit in der äußeren Form und im inneren Bau dennoch so zahlreiche und wichtige gemeinsame Grundzüge, daß wir an ihrer Blutsverwandtschaft nicht zweifeln können. Dasselbe gilt auch von den sechs großen Hauptklassen, welche die neuere Botanik im Pflanzenreiche unterscheidet, nämlich 1. die Blumenpflanzen (Phanerogamae); 2. die Farne (Filicinae); 3. die Mose (Muscinae); 4. die Flechten (Lichenes); 5. die Pilze (Fungi) und 6. die Tange (Algae). Die letzten drei Gruppen zeigen selbst wiederum unter sich so nahe Beziehungen, daß man sie als Thalluspflanzen (Thallophyta) den drei erstn Hauptklassen gegenüber stellen, und somit die Zahl der Phylen oder Hauptgruppen des Pflanzenreichs auf vier beschränken könnte. Auch Mose und Farne könnte man als Prothalluspflanzen (Prothallophyta) zusammenfassen und dadurch die Zahl der Pflanzenstämme auf drei erniedrigen: Blumenpflanzen, Prothalluspflanzen und Thalluspflanzen.
Nun sprechen aber sehr gewichtige Thatsachen der Anatomie und der Entwickelungsgeschichte sowohl im Thierreich als im Pflanzenreich für die Vermuthung, daß auch diese wenigen Hauptklassen oder Stämme noch an ihrer Wurzel zusammenhängen, d. h. daß ihre niedersten und ältesten Stammformen unter sich wiederum blutsverwandt sind. Ja bei weiter gehender Untersuchung werden wir noch einen Schritt weiter und zu Darwin's Annahme hingedrängt, daß auch die beiden Stammbäume des Thier- und Pflanzenreichs an ihrer tiefsten Wurzel zusammenhängen, daß auch die niedersten und ältesten Thiere und Pflanzen von einem einzigen gemeinsamen Urwesen abstammen. Natürlich könnte nach unserer Ansicht dieser gemeinsame Urorganismus nur ein durch Urzeugung entstandenes Moner sein.
Vorsichtiger werden wir vorläufig jedenfalls verfahren, wenn wir diesen letzten Schritt noch vermeiden, und wahre Blutsverwandtschaft nur innerhalb jedes Stammes oder Phylum annehmen, wo sie durch die Thatsachen der vergleichenden Anatomie, Ontogenie und Phylogenie unzweifelhaft sicher gestellt wird. Aber schon jetzt können wir bei dieser Gelegenheit darauf hinweisen, daß zwei verschiedene Grundformen der genealogischen Hypothesen möglich sind, und daß alle verschiedenen Untersuchungen der Descendenztheorie über den Ursprung der organischen Formengruppen sich künftig entweder mehr in der einen oder mehr in der anderen von diesen beiden Richtungen bewegen werden. Die einheitliche (einstämmige oder monophyletische) Abstammungshypothese wird bestrebt sein, den ersten Ursprung sowohl aller einzelnen Organismengruppen als auch der Gesammtheit derselben auf eine einzige gemeinsame, durch Urzeugung entstandene Monerenart zurückzuführen. Die vielheitliche (vielstämmige oder polyphyletische) Descendenztheorie dagegen wird annehmen, daß mehrere verschiedene Monerenarten durch Urzeugung entstanden sind, und daß diese mehreren verschiedenen Hauptklassen (Stämmen oder Phylen) den Ursprung gegen haben. Im Grunde ist der scheinbar sehr bedeutende Gegensatz zwischen diesen beiden Hypothesen von sehr geringer Wichtigkeit. Denn beide, sowohl die einheitliche oder monophyletische, als die vielheitliche oder polyphyletische Descendenzhypothese, müssen nothwendig auf Moneren als auf die älteste Wurzel des einen oder der vielen organischen Stämme zurückgehen. Da aber der ganze Körper aller Moneren nur aus einer einfachen, stucturlosen und formlosen Masse, einer einzigen eiweißartigen Kohlenstoffverbindung besteht, so können die Unterschiede der verschiedenen Moneren nur chemischer Natur sein und nur in einer verschiedenen atomistischen Zusammensetzung jener schleimartigen Eiweißverbindung bestehen. Diese feinen und verwickelten Mischungsverschiedenheiten der unendlich mannichfaltig zusammengesetzten Eiweißverbindungen sind aber vorläufig für die rohen und groben Erkenntnißmittel des Menschen gar nicht erkennbar, und daher auch für unsere vorliegende Aufgabe zunächst von weiter keinem Interesse.
Die Frage nach dem einheitlichen oder vielheitlichen Ursprung wird sich auch innerhalb jedes einzelnen Stammes immer wiederholen, wo es sich um den Ursprung einer kleineren oder größeren Gruppe handelt. Im Pflanzenreiche z. b. werden die einen Botaniker mehr geneigt sein, die sämmtlichen Blumenpflanzen von einer einzigen Farnform abzuleiten, während die anderen die Vorstellung vorziehen werden, daß mehrere verschiedene Phanerogamengruppen aus mehreren Farngruppen hervorgegangen sind. Ebenso werden im Thierreich die einen Zoologen mehr zu Gunsten der Annahme sein, daß sämmtliche placentalen Säugethiere von einer einzigen Beutelthierform abstammen, die anderen dagegen mehr zu Gunsten der entgegensetzten Annahme, daß mehrere verschiedene Gruppen von Placentalthieren aus mehreren verschiedenen Beutelthiergruppen hervorgegangen sind. Was das Menschengeschlecht selbst betrifft, so werden die Einen den Ursprung derselben aus einer einzigen Affenform vorziehen, während die Anderen mehr zu der Vorstellung neigen werden, daß mehrere verschiedene Menschenarten unabhängig von einander aus mehreren verschiedenen Affenarten entstanden sind. Ohne uns hier schon bestimmt für die eine oder die andere Auffassung auszusprechen, wollen wir dennoch die Bemerkung nicht unterdrücken, daß im Allgemeinen die einstämmigen oder monophyletischen Descendenzhypothesen den Vorzug vor den vielstämmigen oder polyphyletischen Abstammungshypothesen verdienen, und zwar vorläufig schon aus dem einfachen Grunde, weil sie die unendlich schwierige Aufgabe der Stammbaumconstructionen in hohem Grade erleichtern. Es ist jedoch möglich, daß die entwickeltere Descendenztheorie der Zukunft den polyphyletischen Ursprung insbesondere für viele niedere und unvollkommene Gruppen der beiden organischen Reiche nachweisen wird. Gegenwärtig aber würden wir, wollten wir denselben verfolgen, jedenfalls in ein unentwirrbares Labyrinth von dunklen und widersprechenden Vermuthungen uns verlieren.
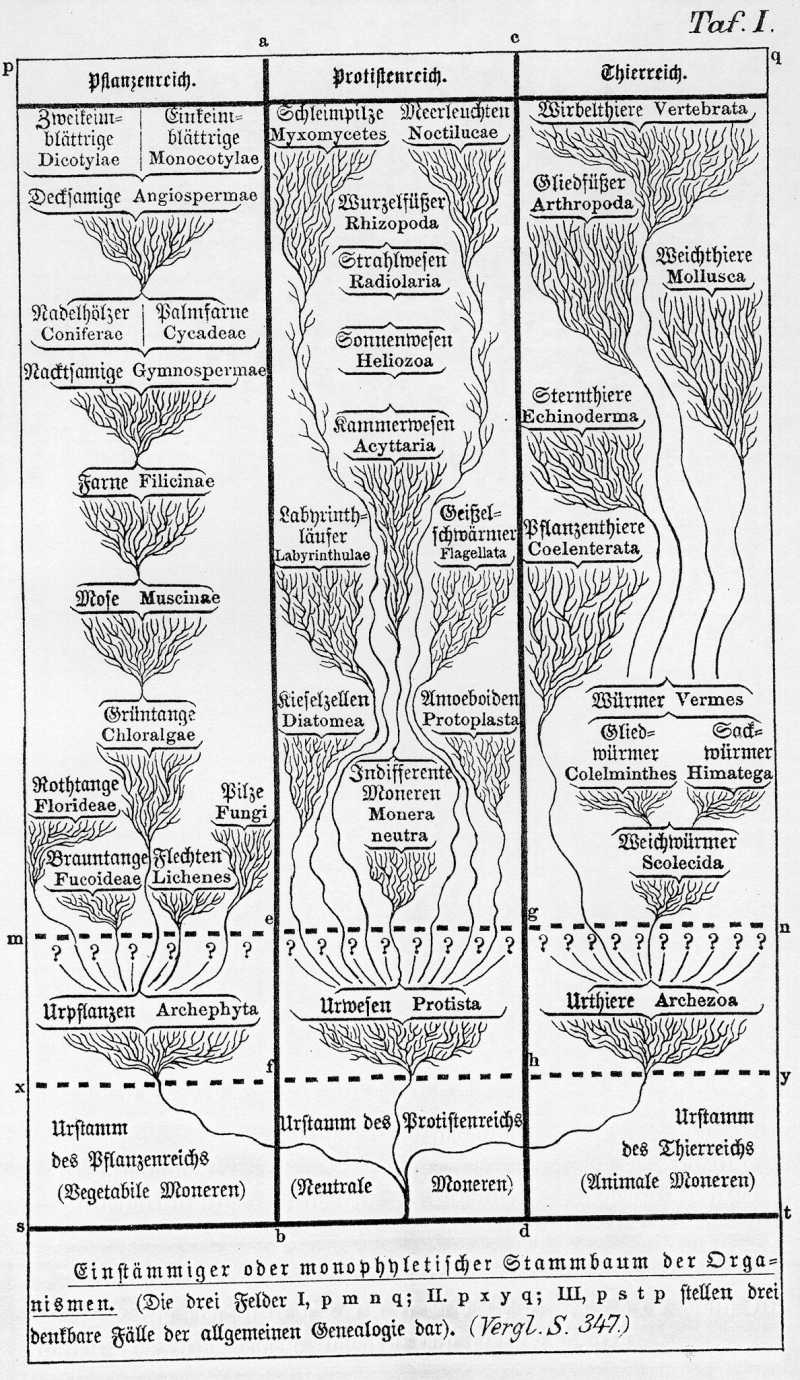
Aus diesem Grunde halte ich es für das Beste, gegenwärtig für das Thierreich einerseits, für das Pflanzenreich andrerseits eine einstämmige der monophyletische Descendenzhypothese anzunehmen, ungefähr in der Form, wie sie auf Taf. II. und III graphisch dargestellt ist. Hiernach würden also die oben genannten sechs Stämme oder Phylen des Thierreichs an ihrer untersten Wurzel zusammenhängen, und ebenso die erwähnten drei bis sechs Hauptklassen oder Phylen des Pflanzenreichs von einer gemeinsamen ältesten Stammform abzuleiten sein. Wie der Zusammenhang dieser Stämme zu denken ist, werde ich in den nächsten Vorträgen erläutern. Zunächst aber müssen wir uns hier noch mit einer sehr merkwürdigen Gruppe von Organismen beschäftigen, welche weder in den Stammbaum des Pflanzenreichs (Taf. II.), noch in den Stammbaum des Thierreichs (Taf. III.) ohne künstlichen Zwang eingereiht werden können. Diese intressanten und wichtigen Organismen sind die Urwesen oder Protisten. (Vergl. Taf. I.).
Sämmtliche Organismen, welche wir als Protisten zusammenfassen, zeigen in ihrer äußeren Form, in ihrem inneren Bau und in ihren gesammten Lebenserscheinungen eine so merkwürdige Mischung von thierischen und pflanzlichen Eigenschaften, daß sie mit klarem Rechte weder dem Thierreiche, noch dem Pflanzenreiche zugetheilt werden können, und daß seit mehr als zwanzig Jahren ein endloser und fruchtloser Streit darüber geführt wird, ob sie in jenes oder in dieses einzuordnen seien.
Die meisten Protisten oder Urwesen sind von so geringer Größe, daß man sie mit bloßem Auge nur schwer oder gar nicht wahrnehmen kann. Daher ist die Mehrzahl derselben erst im Laufe der letzten fünfzig Jahre bekannt geworden, seit man mit Hülfe der verbesserten und allgemein verbreiteten Mikroskope diese winzigen Organismen häufiger beobachtete und genauer untersuchte. Aber sobald man dadurch näher mit ihnen vertraut wurde, erhoben sich auch alsbald endlose Streitigkeiten über ihre eigentliche Natur und ihre Stellung im natürlichen Systeme der Organismen. Viele von diesen zweifelhaften Urwesen wurden von den Botanikern für Thiere, von den Zoologen für Pflanzen erklärt; es wollte sie keiner von Beiden haben. Andere wurden umgekehrt sowohl von den Botanikern für Pflanzen, als von den Zoologen für Thiere erklärt; jeder wollte sie haben. Diese Wiedersprüche sind nicht etwa durch unsere unvollkommene Kenntniß der Protisten, sondern wirklich durch ihre wahre Natur bedingt. In der That zeigen die meisten Protisten eine so bunte Vermischung von mancherlei thierischen und pflanzlichen Charakteren, daß es lediglich der Willkür des einzelnen Beobachters überlassen bleibt, ob er sie dem Thier- oder Pflanzenreich einreihen will. Je nachdem er diese beiden Reiche definirt, je nachdem er diesen oder jenen Charakter als bestimmend für die Thiernatur oder für die Pflanzennatur ansieht, wird er die einzelnen Protistenklassen bald dem Thierreiche bald dem Pflanzenreiche zuertheilen. Diese systematische Schwierigkeit ist aber dadurch zu einem ganz unauflöslichen Knoten geworden, daß alle neueren Untersuchungen über die niedersten Organismen die bisher übliche scharfe Grenze zwischen Thier- und Pflanzenreich völlig verwischt, oder wenigstens dergestalt zerstört haben, daß ihre Wiederherstellung nur mittelst einer ganz künstlichen Definition beider Reiche möglich ist. Aber auch in diese Definition wollen viele Protisten durchaus nicht hineinpassen.
Aus diesen und vielen anderen Gründen ist es jedenfalls, wenigstens vorläufig das Beste, die zweifelhaften Zwitterwesen sowohl aus dem Thierreiche als aus dem Pflanzenreiche auszuweisen, und in einem zwischen beiden mitten innestehenden dritten organischen Reiche zu vereinigen. Dieses vermittelnde Zwischenreich habe ich als Reich der Urwesen (Protista) in meiner allgemeinen Anatomie (im zweiten Bande der generellen Morphologie) ausführlich begründet (Gen. Morph. I, S. 191-238). In meiner Monographie der Moneren15) habe ich kürzlich dasselbe in etwas veränderter Begrenzung und in schärferer Definition erläutert. Als selbstständige Klassen des Protistenreichs kann man gegenwärtig etwa folgende acht Gruppen ansehen: 1. die noch gegenwärtig lebenden Moneren; 2. die Amoeboiden oder Protoplasten; 3. die Geißelschwärmer oder Flagellaten; 4. die Schleimpilze oder Myxomyceten; 5. die Labyrinthläufer oder Labyrinthuleen; 6. die Kieselzellen oder Diatomeen; 7. die Meerleuchten oder Noctilucen; 8. die Wurzelfüßer oder Rhizopoden.
Wahrscheinlich wird die Anzahl dieser Protistenklassen durch die fortschreitenden Unterschungen über die Ontogenie der einfachsten Lebensformen, die erst seit kurzer Zeit mit größerem Eifer betrieben werden, in Zukunft noch beträchtlich vermehrt werden. Mit den meisten der genannten Klassen ist man erst in den letzten zehn Jahren genauer bekannt geworden. Wahrscheinlich sind auch sehr zahlreiche Protistengruppen in früheren Perioden ausgestorben, ohne uns bei ihrer größtentheils sehr weichen Körperbeschaffenheit fossile Reste hinterlassen zu haben. Von den jetzt noch lebenden niedersten Organismengruppen könnte man dem Protistenreiche auch noch drei andere Klassen anschließen, nämlich einerseits 9. die Phykochromalgen oder Phykochromaceen und 10. die Pilze oder Fungen; andrerseits 11. die Schwämme oder Spongien. Indessen erscheint es, (für unsere Betrachtung hier wenigstens) vortheilhafter, die letztere Klasse im Thierreich, die beiden ersteren im Pflanzenreiche stehen zu lassen.
Der Stammbaum des Protistenreichs ist noch in tiefes Dunkel gehüllt. Die eigenthümlich Verbindung von thierischen und pflanzlichen Eigenschaften, der indifferente und unbestimmte Charakter ihrer Formverhältnisse und Lebenserscheinungen, dabei andrerseits eine Anzahl von mehreren, ganz eigenthümlichen Merkmalen, welche die meisten der genannten Klassen scharf von den anderen trennen, vereiteln vorläufig noch jeden Versuch, ihre Blutsverwandtschaft untereinander, oder mit den niedersten Thieren einerseits, mit den niedersten Pflanzen andrerseits, bestimmter zu erkennen. Es ist nicht unwahrscheinlich, daß die genannten und noch viele andere uns unbekannte Protistenklassen ganz selbstständig organische Stämme oder Phylen darstellen, deren jeder sich aus einer, vielleicht sogar aus mehreren, durch Urzeugung entstandenen Moneren unabhängig entwickelt hat. Will man dieser vielstämmigen oder polyphyletischen Descendenzhypothese nicht beipflichten, und zieht man die einstämmige oder monophyletische Annahme von der Blutsverwandtschaft aller Organismen vor, so wird man die verschiedenen Protistenklassen als niedere Wurzelschößlinge zu betrachten haben, aus derselben einfachen Monerenwurzel heraussprossend, aus welcher die beiden mächtigen und vielverzweigten Stammbäume einerseits des Thierreichs, andrerseits des Pflanzenreichs entstanden sind (Taf. I.) Bevor ich Ihnen diese schwierige und dunkle Frage näher erläutere, wird es wohl passend sein, noch Einiges über den Inhalt der vorstehend angeführten Protistenklassen und ihre allgemeine Naturgeschichte vorauszuschicken.
Das ich hier wieder mit den merkwürdigen Moneren (Monera) als erster Klasse des Protistenreiches beginne, wird Ihnen vielleicht seltsam vorkommen, da ich ja Moneren als die ältesten Stammformen aller Organismen ohne Ausnahme ansehe. Allein was sollen wir sonst mit den gegenwärtig noch lebenden Moneren anfangen? Wir wissen Nichts von ihrem paläontologischen Ursprung, wir wissen Nichts von irgend welchen Beziehungen derselben zu niederen Thieren oder Pflanzen, wir wissen Nichts von ihrer möglichen Entwicklungsfähigkeit zu höheren Organismen. Das structurlose und homogene Schleimklümpchen, welches ihren ganzen Körper bildet, ist ebenso die älteste und ursprünglichste Grundlage der thierischen wie der pflanzlichen Plastiden. Offenbar würde es daher ebenso willkürlich und grundlos sein, wenn man sie dem Thierreiche, als wenn man sie dem Pflanzenreiche anschließen wollte. Jedenfalls verfahren wir vorläufig am vorsichtigsten und am meisten kritisch, wenn wir die gegenwärtig noch lebenden Moneren, deren Zahl und Verbreitung vielleicht sehr groß ist, als eine ganz besondere und selbstständige Klasse zusammenfassen, welche wir allen übrigen Klassen sowohl des Protistenreichs, als des Pflanzereichs und des Thierreichs gegenüber stellen.
Durch die vollkommene Gleichartigkeit ihrer ganzen eiweißartigen Körpermasse, durch den völligen Mangel einer Zusammensetzung aus ungleichartigen Theilchen schließen sich, rein morphologisch betrachtet, die Moneren näher an die Anorgane als an die Organismen an, und vermitteln offenbar den Uebergang zwischen anorganischer und organischr Körperwelt, wie ihn die Hypothese der Urzeugung annimmt. Da ich Ihnen die Lebenserscheinungen der jetzt noch lebenden Moneren (Protamoeba, Protogenes, Protomyxa etc. ) bereits früher geschildert habe, so verweise ich Sie auf den achten Vortrag (S. 142) und auf meine Monographie der Moneren,15) und wiederhole hier nur als Beispiel die früher gegebene Abbildung der Protamoeba, eines Moneres, welches das süße Wasser bewohnt (Fig. 12). Nicht weniger genealogische Schwierigkieten als die Moneren, bieten uns die Amoeben der Gegenwart, und die ihnen nächstverwandten Organismen (Arcelliden und Gregarinen), welche wir hier als eine zweite Protistenklasse unter dem Namen der Amoeboiden (Protoplasta) zusammenfassen. Man stellt diese Urwesen jetzt gewöhnlich in das Thierreich, ohne daß man eigentlich einsieht, warum? Denn einfache nackte Zellen, d. h. hüllenlose und kernführende Plastiden, kommen eben sowohl bei echten Pflanzen als bei echten Thieren vor. Eigentlich ist jede nackte einfache Zelle, gleichviel ob sie aus dem Thier- oder Pflanzenkörper kömmt, von einer selbstständigen Amoebe nicht zu unterscheiden.
{Abbildung 12 }
Fig. 12 Fortpflanzung eines einfachsten Organismus, eines Moneres, durch Selbsttheilung. A. Das ganze Moner, eine Protamoeba. B. Dieselbe zerfällt durch eine mittlere Einschnürung in zwei Hälften. C. Jede der beiden Hälften hat sich von der anderen getrennt und stellt nun ein selbstständiges Individuum dar. Denn diese letztere ist selbst Nichts weiter als eine einfache Urzelle, ein nacktes Klümpchen von Zellstoff oder Plasma, welches einen Kern enthält. Die Zusammenziehungsfähigkeit oder Contractilität dieses Plasma aber, welche die freie Amoebe im Ausstrecken und Einziehen formwechselnder Fortsätze zeigt, ist eine allgemeine Lebenseigenschaft des organischen Plasma eben sowohl in den thierischen wie in den pflanzlichen Plastiden. Wenn eine frei bewegliche, ihre Form beständig ändernde Amoebe in den Ruhezustand übergeht, so zieht sie sich kugelig zusammen und umgiebt sich mit einer ausgeschwitzten Membran. Dann ist sie der Form nach ebenso wenig von einem thierischen Ei als von einer einfachen kugeligen Pflanzenzelle zu unterscheiden (Fig. 13 A).
Nackte kernhaltige Zellen, gleich den Fig. 13 B abgebildeten, welche in beständigem Wechsel formlose fingerähnliche Fortsätze ausstrecken und wieder einziehen, und welche man deshalb als Amoeben bezeichnet, finden sich vielfach und sehr weit verbreitet im süßen Wasser und im Meere, so sogar auf dem Lande kriechend vor. Dieselben nehmen ihre Nahrung in derselben Weise auf, wie es früher (S. 143) von den Protamoeben beschrieben wurde. Bisweilen kann man ihre Fortpflanzung durch Theilung (Fig. 13 C, D) beobachten, die ich bereits in einem früheren Vortrag Ihnen geschildert habe (S. 145). Viele von diesen formlosen Amoeben sind neuerdings als jugendliche Entwicklungszustände von anderen Protisten (namentlich den Myxomyceten) oder als abgelöste Zellen von niederen Thieren und Pflanzen erkannt worden.
{Figur 13}
Fig. 13. Fortpflanzung eines einzelligen Organismus, einer Amoeba, durch Selbsttheilung. A. Die eingekapselte Amoeba, eine einfache kugelige Zelle, bestehend aus einen Protoplasmaklumpen (b), welcher einen Kern (a) einschließt, und von einer Zellhaut oder Kapsel umgeben ist. B. Die freie Amoeba, welch die Cyste oder Zellhaut gesprengt und verlassen hat. C. Dieselbe beginnt sich zu theilen, indem ihr Kern in zwei Kerne zerfällt und der Zellstoff zwischen beiden sich einschnürt. D. Die Theilung ist vollendet, indem auch der Zellstoff vollständig in zwei Hälften zerfallen ist (Da und Db).
Die farblosen Blutzellen der Thiere z. B., auch die im menschlichen Blute, sind von Amoeben nicht zu unterscheiden, und können gleich diesen feste Körperchen in ihr Inneres aufnehmen, wie ich zuerst durch Fütterung derselben mit feinzertheilten Farbstoffen nachgewiesen habe (Gen. Morph. I, 271). Andere Amoeben dagegen (wie die in Fig. 13 abgebildeten) scheinen selbstständige "gute Arten oder Species" zu sein, indem sie sich viele Generationen hindurch unverändert fortpflanzen. Außer den eigentlichen oder nackten Amoeben (Gymnamoebae) finden wir weitverbreitet, besonders im süßen Wasser, auch beschalte Amoeben (Lepamoebae), deren nackter Plasmaleib theilweis durch eine mehr oder weniger feste Schale (Arcella) oder selbst ein aus Steinchen zusammengeklebtes Gehäuse (Difflugia) geschützt ist. Endlich finden wir im Leibe von vielen niederen Thieren vielfach schmarotzende Amoeben vor (Gregarinae), welche durch Anpassung an das Schmarotzerleben ihren ganzen Plasmakörper mit einer vollständig geschlossenen Haut umhüllt haben.
Die einfachen nackten Amoeben sind für die gesammte Biologie, und insbesondere für die allgemeine Genealogie, nächst den Moneren die wichtigsten von allen Organismen. Denn offenbar entstanden die Amoeben ursprünglich aus einfachen Moneren (Protamoeba) dadurch, daß der erste wichtige Sonderungsvorgang in ihrem homogenen Schleimkörper stattfand, die Differenzierung des inneren Kerns von dem umgebenden Plasma. Dadurch war der große Fortschritt von einer einfachen (kernlosen) Cytode zu einer echten (kernhaltigen) Zelle geschehen (Vergl. Fig. 12 A und Fig. 13 B). Indem einige von diesen Zellen sich frühzeitig durch Ausschwitzung einer erstarrenden Membran abkapselten, bildeten sie die ersten Pflanzenzellen, während andere, nackt bleibende, sich zu den ersten Zellen des Thierkörpers entwickeln konnten. In der Anwesenehit oder dem Mangel einer umhüllenden starren Membran liegt der wichtigste, obwohl keineswegs durchgreifende Unterschied der pflanzlichen und der thierischen Zellen. Indem die Pflanzenzellen sich schon frühzeitig durch Einschließung in ihre starre, dicke und feste Cellulose-Schale abkapseln, gleich der ruhenden Amoebe, Fig. 13 A, bleiben sie selbstständiger und den Einflüssen der Außenwelt weniger zugänglich, als die weichen, meistens nackten oder nur von einer dünnen und biegsamen Haut umhüllten Thierzellen. Daher vermögen aber auch die ersteren nicht so wie die letzteren zur Bildung höherer, zusammengesetzter Gewebstheile, z. B. Nervenfasern, Muskelfasern zusammenzutreten. Zugleich wird sich bei den ältesten einzelligen Organismen schon frühzeitig der wichtigste Unterschied in der thierischen und pflanzlichen Nahrungsaufnahme ausgebildet haben. Die ältesten einzelligen Thiere konnten als nackte Zellen, so gut wie die freien Amoeben (Fig. 13. B) und die farblosen Blutzellen, feste Körperchen in des Innere ihres weichen Lebes aufnehmen, während die ältesten einzelligen Pflanzen, durch ihre Membran abgekapselt, hierzu nicht mehr fähig waren und bloß flüssige Nahrung (mittelst Diffusion) durch dieselbe durchtreten lassen konnten. Nicht minder zweifelhaft als die Natur der Amoeben ist diejenige der Geißelschwärmer (Flagellata), welche wir als eine dritte Klasse des Protistenreiches betrachten. Auch diese zeigt gleich nahe und wichtige Beziehungen zum Pflanzenreich wie zum Thierreich. Einige Flagellaten sind von den frei beweglichen Jugendstadien echter Pflanzen, namentlich den Schwärmsporen vieler Tange, nicht zu unterscheiden, während andere sich unmittelbar den echten Thieren, und zwar den bewimpterten Infusorien (Ciliata) anschließen. Die Geißelschwärmer sind einfache Zellen, welche entweder einzeln oder zu Colonien vereinigt im süßen und salzigen Wasser leben. Ihr charakteristischer Körper ist ein sehr beweglicher, einfacher oder mehrfacher, peitschenförmiger Anhang (Geißel oder Flagellum), mittelst dessen sie lebhaft im Wasser umherschwärmen.
Die Klasse zerfällt in zwei Ordnungen. Bei den bewimperten Geißelschwärmern (Cilioflagellata) ist außer der langen Geißel auch noch ein Kranz von kurzen Wimpern vorhanden, welcher den unbewimperten Geißelschwärmern (Nudoflagellata) fehlt. Zu den ersteren gehören namentlich die kieselhaltigen gelben Peridinien, welche sich an dem Leuchtes des Meeres stark beteiligen, zu den letzteren die grünen Euglenen, welche oft durch ihre ungeheuren Individuenmassen unsere Teiche im Frühjahr ganz grün färben.
Eine vierte Protistenklasse bilden die merkwürdigen Schleimpilze (Myxomycetes). Diese galten früher allgemein für Pflanzen, für echte Pilze, bis vor neun Jahren der Botaniker de Bary durch Entdeckung ihrer Ontogenie nachwies, daß dieselben gänzlich von den Pilzen verschieden, und eher als niedere Thiere zu betrachten seien. Allerdings ist der reife Fruchtkörper derselben eine rundliche, oft mehrere Zoll große, mit feinem Sporenpulver und weichen Flocken gefüllte Blase, wie bei den bekannten Bovisten oder Bauchpilzen (Gastromycetes). Allein aus den Keimkörnern oder Sporen derselben kommen nicht die charakteristischen Fadenzellen oder Hyphen der echten Pilze hervor, sondern nackte Zellen, welche anfangs in Form von Geißelschwärmern umherschwimmen, später nach Art der Amoeben umherkriechen und endlich mit anderen Ihresgleichen zu großen Schleimkörpern oder "Plasmodien" zusammenfließen, aus denen dann unmittelbar der blasenförmige Fruchtkörper entsteht. Wahrscheinlich kennen Sie Alle eines von jenen Plasmodien, dasjenige von Aethalium septicum, welches im Sommer als sogenannte "Lohblüthe" in Form einer schöngelben, oft mehrere Fuß breiten, salbenartigen Schleimmasse netzförmig die Lohhaufen und Lohbeete der Gerber durchzieht. Die schleimigen frei kriechenden Jugendzustände dieser Myxomyceten, welche meistens auf faulenden Pflanzenstoffen, Baumrinden u. s. w. in feuchten Wäldern leben, werden mit gleichem Recht oder Unrecht von den Zoologen für Thiere, wie die reifen und ruhenden blasenförmigen Fruchtzustände von den Botanikers für Pflanzen erklärt.
Nicht weniger räthselhafter Natur sind ebenfalls die Protisten der fünften Klasse, die Labyrinthläufer (Labyrithuleae), welche erst kürzlich von Cienkowsky an Pfählen im Seewasser entdeckt wurden. Es sind spindelförmige, meistens dottergelb gefärbte Zellen, welche bald in dichten Haufen zu Klumpen vereinigt sitzen, bald in höchst eigenthümlicher Weise sich umherbewegen. Sie bilden dann in noch unerklärter Weise ein netzförmiges Gerüst von labyrinthisch verschlungenen Strängen, und in der starren "Fadenbahn" dieses Gerüstes rutschen sie umher. Der Gestalt nach würde man die Zellen der Labyrinthuleen für einfachste Pflanzen, der Bewegung nach für einfachste Thiere halten. In der That sind sie weder Thiere noch Pflanzen.
Den Labyrinthuleen verwandt nächstverwandt sind die Kieselzellen (Diatomeae), eine sechste Protistenklasse. Diese Urwesen, welche meistens für Pflanzen, aber von einigen berühmten Naturforschern noch heute für Thiere gehalten werden, bevölkern in ungeheuren Massen und in einer unendlichen Mannichfaltigkeit der zierlichsten Formen das Meer und die süßen Gewässer. Meist sind es mikroskopisch kleine Zellen, welche entweder einzeln oder in großer Menge vereinigt leben, und entweder festgewachsen sind oder sich in eigenthümlicher Weise rutschend, schwimmend oder kriechend, umherbewegen. Ihr weicher Zellenleib, der durch einen charakteristischen Farbstoff bräunlich gelb gefärbt ist, wird stets von einer festen und starren Kieselschale umschlossen, welche die zierlichsten und mannichfaltigsten Formen besitzt. Diese Kieselhülle ist nur durch eine oder ein paare Spalten nach außen geöffnet und läßt dadurch den eingeschlossenen weichen Plasmaleib mit der Außenwelt communiciren. Die Kieselschalen finden sich massenhaft versteinert vor und setzen manche Gesteine, z. B. den Biliner Polirschiefer, das schwedische Bergmehl u. s. w. vorwiegend zusammen. Eine eigene, siebente Protistenklasse bilden die Meerleuchten (Noctilucae). Es sind kleine, weiche, schleimige Bläschen, von der Form einer Pfirsich. Sie haben gewöhnlich nur etwa eine halbe Linie oder einen Millimeter Durchmesser, bedecken aber die Meeresoberfläche oft in so erstaunlichen Massen, daß sie in meilenweiter Ausdehnung eine mehr als zolldicke Schleimschicht auf der Oberfläche bilden. Sie gehören neben den obenerwähnten Peridinien, und neben vielen niederen Seethieren (besonders Medusen und Krebsen) zu den wesentlichesten Ursachen des Meerleuchtens, indem sie im Dunkeln einen phosphorischen Glanz austrahlen. Trotzdem sie in so ungeheuren Massen in der Nordsee, im Mittelmeere u. s. w. vorkommen, kennen wir dennoch die Naturgeschichte der Noctiluken nur sehr unvollständig. Es ist möglich, daß sie den Pflanzen näher als den Thieren verwandt sind, obwohl die meisten Naturforscher sie gegenwärtig zu den Thieren zählen. Wahrscheinlich sind es neutrale Protisten.
Ebenso zweifelhaft ist auch die Natur der achten und letzten Klasse des Protistenreichs, der Wurzelfüßer (Rhizopoda). Diese merkwürdigen Organismen bevölkern das Meer seit den ältesten Zeiten der organischen Erdgeschichte in einer außerordentlichen Formenmannichfaltigkeit, theils auf dem Meeresboden kriechend, theils an der Oberfläche schwimmend. Nur sehr wenige leben im sußen Wasser (Gromia, Actinosphaerium). Die meisten besitzen feste, als Kalkerde oder Kieselerde bestehende und höchst zierlich zusammengesetzte Schalen, welche in versteinerten Zustande sich vortrefflich erhalten. Oft sind dieselben sehr klein und häufig für das bloße Auge kaum oder gar nicht sichtbar sind. Nur wenige erreichen einen Durchmesser von einigen Linien oder selbst von ein paar Zollen. Ihren Namen führt die ganze Klasse davon, daß ihr nackter schleimiger Leib an der ganzen Oberfläche tausende von äußerst feinen Schleimfäden ausstrahlt, falschen Füßchen, Scheinfüßchen oder Pseudopodien, welche sich wurzelförmig verästeln, netzförmig verbinden, und in beständigem Formwechsel gleich der einfacheren Schleimfüßchen der Amoeboiden oder Protoplasten befindlich sind. Diese veränderlichen Scheinfüßchen dienen sowohl zur Ortsbewegung, als zur Nahrungsaufnahme.
Die Klasse der Wurzelfüßer zerfällt in drei verschiedene Legionen, die Kammerwesen oder Acyttarien, die Sonnenwesen oder Heliozoen und die Strahlwesen oder Radiolarien. Die erste und niederste von diesen drei Legionen bilden die Kammerwesen (Acyttaria). Hier besteht nämlich der ganze weiche Leib noch aus einfachem schleimigem Zellstoff oder Plasma, das noch nicht in Zellen differenzirt ist. Allein trotz dieser höchst primitiven Leibesbeschaffenheit schwitzen die Kammerwesen dennoch eine meistens feste, aus Kalkerde bestehende Schale aus, welche eine große Mannichfaltigkeit zierlicher Formbildung zeigt. Bei den älteren und einfacheren Acyttarien ist diese Schale eine einfache, glockenförmige, röhrenförmige oder schneckenhausförmige Kammer, aus deren Mündung ein Bündel von Schleimfäden hervortritt. Im Gegensatz zu diesen Einkammerwesen (Monothalamia) besitzen die Vielkammerwesen (Polythalamia), zu denen die große Mehrzahl der Acyttarien gehört, ein Gehäuse, welches aus zahlreichen Kammern in sehr künstlicher Weise zusammengesetzt ist. Bald liegen diese Kammern in einer Reihe hinter einander, bald in concentrischen Kreisen oder Spiralen ringförmig um einen Mittelpunkt herum, und dann oft in vielen Etagen übereinander, gleich den Logen eines großen Amphitheaters. Diese Bildung besitzen z. B. die Nummuliten, deren linsengroße Kalkschalen, zu Milliarden angehäuft, an der Mittelmeerküste ganze Gebirge zusammensetzen. Die Steine, aus denen die egyptischen Pyramiden aufgebaut sind, bestehen aus solchem Nummulitenkalk. In den meisten Fällen sind die Schalenkammern der Polythalamien in einer Spirallinie um einander gewunden. Die Kammern stehen mit einander durch Gänge und Thüren in Verbindung, gleich den Zimmern eines großen Palastes, und sind nach außen gewöhnlich durch zahlreiche kleine Fenster geöffnet, aus denen der schleimige Körper formwechselnde Scheinfüßchen ausstrecken kann. Und dennoch, trotz des außerordentlich verwickelten und zierlichen Baues dieser Kalkpaläste, trotz der unendlichen Mannichfaltigkeit in dem Bau und der Verzierung seiner zahlreichen Kammern, trotz der Regelmäßigkeit und Eleganz ihrer Ausführung, ist dieser ganze künstliche Palast das augeschwitzte Product einer vollkommen formlosen und structurlosen Schleimmasse! Fürwahr, wenn nicht schon die ganze neuere Anatomie der thierischen und pflanzlichen Gewebe unsere Plastidentheorie stützte, wenn nicht alle allgemeinen Resultate derselben übereinstimmend bekräftigten, daß das ganze Wunder der Lebenserscheinungen und Lebensformen auf die active Thätigkeit der formlosen Eiweißverbindungen des Plasma zurückzuführen ist, die Polythalamien allein schon müßten unserer Theorie den Sieg verleihen. Denn hier können wir jeden Augenblick die wunderbare, aber unleugbare und zuerst von Dujardin und Max Schultze24) festgestellte Thatsache durch das Mikroskop nachweisen, daß der formlose Schleim des weichen Plasmakörpers, dieser wahre "Lebensstoff", die zierlichsten, regelmäßigsten und verwickeltesten Bildungen auszuscheiden vermag. Dadurch lernen wir verstehen, wie derselbe "Urschleim", dasselbe Protoplasma, im Körper der Thiere und Pflanzen die verschiedensten Zellenformen erzeugen kann.
Von ganz besonderem Interesse ist es noch, daß zu den Polythalamien auch der älteste Organismus gehört, dessen Reste uns in versteinertem Zustande erhalten sind. Dies ist das vorher bereits erwähnte "kanadische Morgenwesen", Eozoon canadense, welches vor wenigen Jahren in der Ottawaformation (in den tiefsten Schichten des laurentischen Systems) am Ottawaflusse in Canada gefunden worden ist. In der That, durften wir überhaupt erwarten, in diesen ältesten Ablagerungen der Primordialzeit noch organische Reste zu finden, so konnten wir vor Allen auf diese einfachsten und doch mit einer festen Schale bedeckten Protisten hoffen, in deren Organisation der Unterschied zwischen Thier und Pflanze noch nicht ausgeprägt ist.
Von der zweiten Klasse der Wurzelfüßer, von den Sonnenwesen (Heliozoa), kennen wir nur eine einzige Art, das sogenannte "Sonnenthierchen", welches in unseren süßen Gewässern sehr häufig ist. Schon im vorigen Jahrhundert wurde dasselbe von Pastor Eichhorn in Danzig beobachtet und nach ihm Actinophaerium Eichhornii getauft. Es erscheint dem bloßen Auge als ein gallertiges graues Schleimkügelchen von der Größe eines Stecknadelkopfes. Unter dem Mikroskope sieht man Hunderte oder Tausende feiner Schleimfäden von dem centralen Plasmakörper ausstrahlen, und bemerkt, das seine innere zellige Markschicht von der äußeren blasigen Rindenschicht verschieden ist. Dadurch erhebt sich das kleine Sonnenwesen, trotz des Mangels einer Schale, bereits über die structurlosen Acyttarien und bildet den Uebergang von diesen zu den Radiolarien.
Die Strahlwesen (Radiolaria) bilden die dritte und letzte Klasse der Rhizopoden. In ihren niederen Formen schließen sie sich eng an die Sonnenwesen und Kammerwesen an, während sie sich in ihren höheren Formen weit über diese erheben. Von Beiden unterscheiden sie sich wesentlich dadurch, daß der centrale Theil des Körpers aus vielen Zellen zusammengesetzt und von einer festen Membran umhüllt ist. Diese geschlossene, meistens kugelige "Centralkapsel" ist in eine schleimige Plasmaschicht eingehüllt, von welcher überall Tausende von höchst feinen Fäden, die verästelten und zusammenfließenden Scheinfüßchen, ausstrahlen. Dazwischen sind zahlreiche gelbe Zellen von räthselhafter Bedeutung zerstreut.
Die meisten Radiolarien zeichnen sich durch ein sehr entwickeltes Skelet aus, welches aus Kieselerde besteht, und eine wunderbare Fülle der zierlichsten und seltsamsten Formen zeigt. Bald bildet dieses Kieselskelet eine einfache Gitterkugel (Fig. 14 s), bald ein künstliches System von mehreren concentrischen Gitterkugeln, welche in einander geschachtelt und durch radiale Stäbe verbunden sind. Meistens strahlen zierliche, oft baumförmig verzweigte Stacheln von der Oberfläche der Kugeln aus. Anderemale besteht das ganze Skelet bloß aus einem Kieselstern und ist dann meistens aus zwanzig, nach einem bestimmten mathematischen Gesetze vertheilten und in einem gemeinsamen Mittelpunkte vereinigten Stacheln zusammengesetzt. Bei noch anderen Radiolarien bildet das Skelet zierliche vielkammerige Gehäuse wie bei den Polythalamien. Es giebt wohl keine andere Gruppe von Organismen, welche eine solche Fülle der verschiedenartigsten Grundformen und eine so geometrische Regelmäßigkeit, verbunden mit der zierlichsten Architektonik, in ihren Skeletbildungen entwickelte. Die meisten der bis jetzt bekannt gewordenen habe ich in dem Atlas abgebildet, der meine Monographie der Radiolarien begleitet23). Hier gebe ich Ihnen als Beispiel nur die Abbildung von einer der einfachsten Gestalten der Cyrtidosphaera echnoides von Nizza25).
{ Figur 14..}
Fig. 14. Cyrtidosphaera echinoides, 400mal vergrößert. c. Kugelige Centralkapsel. s. Gitterförmig durchbrochene Kieselschale. a. Radiale Stacheln, welche von derselben ausstrahlen. p. Pseudopodien oder Scheinfüßchen, welche von der die Centralkapsel umgebenden Schleimhülle ausstrahlen. l. Gelbe kugelige Zellen, welche dazwischen zerstreut sind.
Das Skelet besteht hier bloß aus einer einfachen Gitterkugel (s), welche kurze radiale Stacheln (a) trägt, und welche die Centralkapsel (c) locker umschließt. Von der Schleimhülle, die letztere umgiebt, strahlen sehr zahlreiche und feine Scheinfüßchen (p) aus, welche links zum Theil zurückgezogen und in einen klumpige Schleimmasse verschmolzen sind. Dazwischen sind viele gelbe Zellen (l) zerstreut.
Während die Acyttarien meistens nur auf dem Grunde des Meeres leben, auf Steinen und Seepflanzen, zwischen Sand und Schlamm mittelst ihrer Scheinfüßchen umherkriechend, schwimmen dagegen die Radiolarien meistens an der Oberfläche des Meeres, mit rings ausgestreckten Pseudopodien flottirend. Sie finden sich hier in ungeheuren Mengen beisammen, sind aber meistens so klein, daß man sie bis vor zwanzig Jahren fast völlig übersah und erst seit zehn Jahren genauer kennen lernte. Fast nur diejenigen Radiolarien, welche in Gesellschaften beisammen leben (Polycyttarien) bilden Gallertklumpen von einigen Linien Durchmesser.
Dagegen die meisten einzeln lebenden (Monocyttarien) kann man mit bloßen Auge nicht sehen. Trotzdem finden sich ihre versteinerten Schalen in solchen Massen angehäuft, daß sie an manchen Stellen ganze Berge zusammensetzen, z. B. die Nikobareninseln bei Hinterindien und die Insel Barbados in den Antillen. Da die Meisten von Ihnen mit den eben angeführten acht Protistenklassen vermuthlich nur sehr wenig oder vielleicht gar nicht genauer bekannt sein werden, so will ich jetzt zunächst noch einiges Allgemeine über ihre Naturgeschichte bemerken. Die große Mehrzahl aller Protisten lebt im Meere, theils freischwimmend an der Oberfläche der See, theils auf dem Meeresboden kriechend, oder an Steinen, Muscheln, Pflanzen u. s. w. festgewachsen. Sehr viele Arten von Protisten leben auch im süßen Wasser, aber nur eine sehr geringe Anzahl auf dem festen Lande (z. B. die Myxomyceten, einige Protoplasten). Die meisten können nur durch das Mikroskop wahrgenommen werden, ausgenommen, wenn sie zu Millionen von Individuen zusammengehäuft vorkommen. Nur Wenige erreichen einen Durchmesser von mehreren Linien oder selbst einigen Zollen. Was ihnen aber an Körpergröße abgeht, ersetzen sie durch die Production erstaunlicher Massen von Individuen, und greifen dadurch oft sehr bedeutend in die Oekonomie der Natur ein. Die unverweslichen Ueberreste der gestorbenen Protisten, wie die Kieselschalen der Diatomeen und Radiolarien, die Kalkschalen der Acyttarien, setzen oft dicke Gebirgsschichten zusammen.
In ihren Lebenserscheinungen, insbesondere in Bezug auf Ernährung und Fortpflanzung, schließen sich die einen Protisten mehr den Pflanzen, die anderen mehr den Thieren an. Die Nahrungsaufnahme sowohl als der Stoffwechesel gleicht bald mehr denjenigen der niederen Thiere, bald mehr denjenigen der niederen Pflanzen. Die meisten Protisten aber zeigen gerade hierin eine merkwürdige Mittelstellung zwischen beiden Reichen. Freie Ortsbewegung kommt vielen Protisten zu, während sie anderen fehlt; allein hierin liegt gar kein entscheidender Charakter, da wir auch unzweifelhafte Thiere kennen, denen die freie Ortsbewegung ganz abgeht, und echte Pflanzen, welche dieselbe besitzen.
Eine Seele besitzen alle Protisten, so gut wie alle Thiere und Pflanzen. Die Seele scheint bei vielen Protisten sehr zarter Empfindung fähig zu sein; wenigstens sind dieselben höchst reizbar. Dagegen scheint der Wille bei den meisten sehr schwach entwickelt zu sein, und ob irgend ein Protist selbstständiges Denkvermögen besitzt, ist sehr zweifelhaft. Allein das Denkvermögen fehlt in gleichem Grade auch vielen niederen Thieren, während viele von den höheren Thieren ebenso klar und oft folgerichtiger als viele niedere Menschen denken.
Der wichtigste physiologische Charakter des Protistenreichs liegt in der ausschließlich ungeschlechtlichen Fortpflanzung aller hierher gehörigen Organismen. Die höheren Thiere und Pflanzen pflanzen sich fast ausschließlich nur auf geschlechtlichem Wege fort. Die niederen Thiere und Pflanzen vermehren sich zwar auch vielfach auf ungeschlechtlichem Wege, durch Theilung, Knospenbildung, Keimbildung u. s. w. Allein daneben findet sich bei denselben doch fast immer noch die geschlechtliche Fortpflanzung, oft mit ersterer regelmäßig in Generationen abwechselnd (Metagenesis S. 88). Sämmtliche Protisten dagegen pflanzen sich ausschließlich nur auf dem ungeschlechtlichen Wege fort und der Gegensatz der beiden Geschlechter ist bei ihnen überhaupt noch nicht durch Differenzirung entstanden. Es giebt weder männliche noch weibliche Protisten.
Wie die Protisten in ihren Lebenserscheinungen zwischen Thieren und Pflanzen (und zwar vorzüglich zwischen den niedersten Formen derselben) mitten inne stehen, so gilt dasselbe auch von der chemischen Zusammensetzung ihres Körpers. Einer der wichtigsten Unterschiede in der chemischen Zusammensetzung des Thier- und Pflanzenkörpers besteht in ihrer charakteristischen Skeletbildung. Das Skelet oder das feste Gerüste des Körpers besteht bei den meisten echten Pflanzen aus der stickstofffreien Cellulose, welche ein Ausschwitzungsprodukt des stickstoffhaltigen Zellstoffs oder Protoplasma ist. Bei den meisten echten Thieren dagegen besteht das Skelet gewöhnlich entweder aus stickstoffhaltigen Verbindungen (Chitin u. s. w.), oder aus Kalkerde. In dieser Beziehung verhalten sich die einen Protisten mehr wie Pflanzen, die anderen mehr wie Thiere. Bei Vielen ist das Skelet vorzugsweise oder ganz aus Kieselerde gebildet, welche sowohl im Thier- als im Pflanzenkörper vorkommt. Der active Lebensstoff ist aber in allen Fällen immer das schleimige Protoplasma.
In Bezug auf die Formbildung der Protisten ist insbesondere hervorzuheben, daß die Individualität ihres Körpers fast immer auf einer außerordentlich tiefen Stufe der Entwickelung stehen bleibt. Sehr viele Protisten bleiben zeitlebens einfache Plastiden oder Individuen erster Ordnung. Andere bilden zwar durch Vereinigung von mehreren Individuen Colonien oder Staaten von Plastiden. Allein auch diese höheren Individuen zweiter Ordnung bleiben meistens auf einer sehr tiefen Ausbildungsstufe stehen. Die Bürger dieser Plastidengemeinden bleiben sehr gleichartig, gehen nur in sehr geringem Grade Arbeitstheilung ein, und vermögen daher ebenso wenig ihren staatlichen Organismus zu höheren Leistungen zu befähigen, als etwa die Wilden Neuhollands dies im Stande sind. Der Zusammenhang der Plastiden bleibt auch meistens sehr locker, und jede einzelne bewahrt in hohem Maße ihre individuelle Selbstständigkeit. Individualitäten höherer (dritter bis sechster) Ordnung, wie sie im Thier- und Pflanzenreiche sehr allgemein ausgebildet sind, finden wir unter den Protisten nur in geringer Verbreitung entwickelt.
Ein zweiter Formcharakter, welcher nächst der niederern Individualitätsstufe die Protisten besonders auszeichnet, ist der niedere Ausbildungsgrad ihrer stereometrischen Grundform. Wie ich in meiner Grundformenlehre (im vierten Buche der generellen Morphologie) gezeigt habe, ist bei den meisten Organismen sowohl in der Gesammtbildung des Körpers als in der Form der einzelnen Theile eine bestimmte geometrische Grundform nachzuweisen. Diese ideale Grundform, welche durch die Zahl, Lagerung, Verbindung und Differenzirung der zusammensetzenden Theile bestimmt ist, verhält sich zu der realen organischen Form ganz ähnlich, wie sich die ideale geometrische Grundform der Krystalle zu ihrer unvollkommenen realen Form verhält. Bei den meisten Körprn und Körpertheilen von Thieren und Pflanzen ist diese Grundform eine Pyramide, und zwar bei den sogenannten "strahlig- regulären" Formen eine reguläre Pyramide, bei den höher differenzirten, sogenannten "bilateral-symmetrischen Formen eine irreguläre Pyramide (Vergl. die Tabellen S. 556-558 im zweiten Bande der gen. Morph.). Bei den Protisten ist diese Pyramidenform, welche im Thier- und Pflanzenreiche vorherrscht, im Ganzen selten, und statt dessen ist die Form entweder ganz unregelmäßig (amorph oder irregulär) oder es ist die Grundform eine einfachere reguläre geometische Form, insbesondere sehr häufig die Kugel, der Cylinder, das Ellipsoid, das Sphäroid, der Doppelkegel, der Kegel, das reguläre Vieleck (Tetraeder, Hexaeder, Octaeder, Dodekaeder, Ocosaeder) u. s. w. Alle diese niederen und unvollkommenen Grundformen des promorphologischen Systems sind bei den Protisten die vorherrschenden Grundformen. Jedoch kommen daneben bei vielen Protisten auch noch die höheren regulären und bilateralen Grundformen vor, welche im Thier- und Pflanzenreich herrschend sind. Auch in dieser Hinscht schließen sich oft von nächstverwandten Protisten die einen (z. B. die Acyttarien) mehr den Thieren, die anderen (z. B. die Radiolarien) mehr den Pflanzen an.
Was nun die paläontologische Entwickelung des Protistenreichs betrifft, so kann man sich darüber sehr verschiedene, aber immer nur sehr unsichere genealogische Hypothesen machen. Vielleicht sind die einzelnen Klassen desselben selbstständige Stämme oder Phylen, die sich sowohl unabhängig von einander als von dem Thierreich und von dem Pflanzenreiche entwickelt haben. Dies gilt sowohl wenn wir der vielheitlichen (polyphyletischen) als wenn wir der einheitlichen (monophyletischen) Descendenztheorie folgen. Selbst wenn wir die letztere vollständig annehmen und für alle Organismen ohne Ausnahme, die jemals auf der Erde gelebt haben und noch jetzt leben, die gemeinsame Abstammung von einer einzigen Monerenform behaupten, selbst in diesem Falle ist der Zusammenhang der Protisten einerseits mit dem Pflanzenstamm, andrerseits mit dem Thierstamm nur ein sehr lockerer. Wir hätten sie dann, wie es auf Taf. I dargestellt ist, als niedere Wurzelschößlinge anzusehen, welche sich unmittelbar aus der Wurzel jenes zweistämmigen organischen Stammbaums entwickelt haben, oder vielleicht als tief unten abgehende Zweige eines gemeinsamen niederen Protistenstammes, welcher in der Mitte zwischen den beiden divergirenden hohen und mächtigen Stämmen des Thier- und Pflanzenreichs aufgeschlossen ist. Die einzelnen Protistenklassen, mögen sie nun an ihrer Wurzel gruppenweise enger zusammenhängen oder nur ein lockeres Büschel von Wurzelschößlingen bilden, würden in diesem Falle, ohne weiter mit den rechts nach dem Thierreiche, links nach dem Pflanzenreiche einseitig abgehenden Organismengruppen Etwas zu thun haben, den ursprünglich einfachen Charakter der gemeinsamen Stammform mehr beibehaltn haben, als es bei den echten Thieren und bei den echten Pflanzen der Fall ist.
Nehmen wir dagegen die vielheitliche oder polyphyletische Descendenzhypothese an, so würden wir uns eine mehr oder minder große Anzahl von organischen Stämmen oder Phylen vorzustellen haben, welche alle neben und unabhängig von einander aus dem gemeinsamen Boden der Urzeugung aufschießen. Es würden dann zahlreiche verschiedene Moneren durch Urzeugung entstanden sein, deren Unterschiede nur in geringen, für uns nicht erkennbaren Differenzen ihrer chemischen Zusammensetzung und in Folge dessen auch ihrer Entwickelungsfähigkeit beruhen. Eine geringe Anzahl von diesen Moneren würden den verschiedenen Hauptklassen des Pflanzenreichs, und ebenso andrerseits eine geringe Anzahl den Hauptklassen des Thierreichs den Ursprung gegeben haben. Zwischen beiden Gruppen von Hauptklassen aber würde sich, unabhängig von diesen wie von jenen, eine größere Anzahl von selbstständigen Stämmen entwickelt haben, die auf einer tieferen Organisationsstufe stehen blieben, und sich weder zu echten Pflanzen, noch zu echten Thieren entwickelten. Selbst wenn man einen ganz selbstständigen Stamm für das Pflanzenreich, einen zweiten für das Thierreich annähme, würde man zwischen beiden noch eine größere Anzahl von selbstständigen Protistenstämmen annehmen können, deren jeder ganz unabhängig von jenen aus einer eigenen archigonen Monerenform sich entwickelt hat. Um sich dieses Verhältniß lebendig vorzustellen, werfen Sie einen Blick auf das nachstehende Schema (S. 347), oder stellen Sie sich die ganze Organismenwelt als eine ungeheure Wiese vor, welche größtentheils verdorrt ist, und auf welcher zwei vielverzweigte Bäume stehen, die ebenfalls größtentheils abgestorben sind. Diese letzteren mögen Ihnen das Thierreich und das Pflanzenreich vorstellen, ihre frischen noch grünenden Zweige die lebenden Thiere und Pflanzen, die verdorrten Zweige mit welkem Laub dagegen die ausgestorbenen Gruppen. Das dürre Gras der Wiese entspricht den wahrscheinlich zahlreichen, ausgestorbenen Protistenstämmen, die wenigen noch grünen Halme dagegen den jetzt noch lebenden.
Für die Annahme, daß wiederholt zu verschiedenen Zeiten Moneren durch Urzeugung entstanden sind, spricht vor Allem die Existenz der gegenwärtig noch lebenden Moneren, die ich Ihnen schon früher geschildert habe. Offenbar legen uns diese die Vermuthung sehr nahe, daß der Proceß der Urzeugung noch immer fortdauert.
Denn wir stehen hier vor folgender Alternative. Entweder haben sich seit der ältesten Primordialzeit diese einfachsten Organismen unverändert erhalten und noch bis auf den heutigen Tag, viele Millionen Jahren hindurch, unentwickelt den Charakter der ersten Moneren beibehalten. Oder dies ist nicht der Fall. Dann müssen sich wiederholt durch Urzeugung solche Moneren gebilet haben, und es ist dann nicht abzusehen, warum dieser Prozeß nicht noch immer fortdauern soll. Wie wir bemerkt haben, ist bisher die Urzeugung durch eine wirkliche Beobachtung noch nicht nachgewiesen, was auch jedenfalls (selbst wenn sie alltäglich stattfände!) sehr schwierig sein würde. Allein widerlegt ist die Urzeugung experimentell eben so wenig und kann sie überhaupt niemals werden. Offenbar erscheint es aber bei denkender Betrachtung viel natürlicher, auch jetzt noch diesen Proceß anzunehmen, als zu denken, daß diese einfachsten Schleimklümpchen seit antelaurentischer Zeit noch keinerlei Organe entwickelt und seit jenen vielen Millionen von Jahren sich ganz oder fast ganz unverändert in ihrer primitiven Urgestalt erhalten haben.
{ Abbildung }
Vielstämmiger oder polyphyletischer Stammbaum der Organismen (im Gegensatz zu dem einstämmigen oder monophyletischen Stammbaum auf Taf. I. Die vielen Linien ohne Bezeichnung (mit einem ?) bedeuten zahlreiche ausgestorbene neutrale Stämme des Protistenreichs, welche sich weder zu Thieren noch zu Pflanzen entwickelt haben. (Vergl. S. 345.)
Schöpfungsperioden und Schöpfungsurkunden

 Stammbaum und Geschichte des Pflanzenreichs
Stammbaum und Geschichte des Pflanzenreichs
Renaissance Forum - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - erstes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Jacob Burckhardt - Die Kultur der Renaissance in Italien. Ein Versuch
Leonardo da Vinci Wissenschaftler - Erfinder - Künstler
Venedig - Eine Liebeserklärung an eine Stadt
William Shakespeare animiert (wahrscheinlich oder zufällig...2/3 zu 1/3) William Turner
Philosophie für Schnelldenker - Besinnliche Philosophie
Startseite - © Copyright 2004- - Susanne Albers - Kiehlufer 125 - D 12059 Berlin